人类的起源与华夏种族的形成-何新博客
一、现代人的起源
(一)中国人的起源问题。中华民族是一个晚近的概念,指现在世界上的全体华人。中华民族的前身是华夏种族。4000年前的夏民族与更早的华胥民族,是华夏民族的前身。关于中国人的起源问题,考古界的争论很多,有些学者认为中国人的祖先是北京猿人,后来又有遗传学家提出中国人起源于非洲。本文对有关理论尝试作一梳理。
(二)现代人的诞生
1、现代人的起源。人类进化的最近一个阶段,是包括现代人在内的智人阶段。可分为早期智人和晚期智人。早期智人生活在距今约25万~4万年前,是旧石器时代中期的古人类。晚期智人在解剖结构上属于现代人,大约是在距今四五万年前开始出现的。在晚期智人阶段,人类的分布范围扩大了,从旧大陆到达了澳洲和美洲。这样,由现有发现的化石来追溯人类进化的各个阶段,已可以循着由近及远的序列,从晚期智人—早期智人—直立人—能人,一直追溯到南方古猿。比南方古猿更早、形态更原始的人类的早期代表,则还在探索之中。
在人类起源的问题上,有两个概念:人科的起源和智人的起源。对于人科的共同祖先约700万年前至500万年前起源于非洲的观点,学术界目前并无太大争议。而对于智人的起源,学术界则存在两种假说。智人是人类进化的最后一个阶段,晚期智人的形态已经与现代人基本一致。目前很多科学家支持“非洲起源说”,即非洲是现代人的故乡;还有少数科学家则支持“多地区进化说”,即认为现代人是在欧亚非各自起源。
1987年,美国科学家华莱士和威尔逊分别带领两个实验室通过检测细胞线粒体内的遗传物质脱氧核糖核酸发现,现代人可以追溯其祖先到大约15万年前住在非洲的一个女人。之后,一部分现代人类开始由非洲大陆向世界其他各洲迁移,并在同一时间由穴居人类演变为真正的现代意义上的人类。这一观点至此成为国际学术界争论的“焦点”。
《科学》杂志上公布的文章,剑桥大学的三位研究人员对现代人类从非洲大陆向世界其他各洲迁移的地理位置和相应的人口基因进行分析研究。结果发现距离埃塞俄比亚(非洲东部国家)的人与埃塞俄比亚人间呈现出很小基因变异性。现代人类开始由非洲大陆向世界其他各洲迁移的路线上,基因多样性的特征表现不明显,没有出现大量的基因中断。这就进一步证明了现代人“非洲起源说”。
2、现代人的形成。现代人,即人类进化史上的最新阶段--晚期智人。对于晚期智人的由来,学术界一直存在争议,而且直到今天也不能说这个问题已经解决。如果我们将西欧典型尼安德特人与克罗马农人,或大荔人与山顶洞人的头骨放在一起作一番比较,就会发现它们之间的差异十分明显。典型尼人的头颅又长又低,两侧突出,脑后枕骨像是一个外伸的凸起,额骨向后倾斜,显得很平坦,眉嵴骨十分厚重。然而克罗马农人的头颅很高,胸后枕骨较圆,头骨两侧较垂直,额部也垂直,显得很高。眼眶上没有粗大的眉嵴。脸部的差异也很明显,尼安德特人脸面向前凸出,长有一个很大的鼻子以及硕大而无下颏的下巴。而克罗马农人脸面平,鼻窄,下巴颏突出,已与我们现代人完全一样。
在19世纪,大部分人类进化学者并不将尼安德特人看作是现代人的直系祖先,因为尼安德特人的体质特征过于类似“猿”。当时的观点是将克罗马农人以外的其他化石人类,如中国猿人、爪哇猿人以及尼安德特人都看作是灭绝的旁支。然而到了本世纪20年代,学术界则开始认为尼安德特人是现代人的直系祖先。
但在30年代,1932年发现的斯埋汉姆头骨和1935年发现的斯旺斯科姆头骨使一些学者认为晚期智人早在尼安德特人之前已经出现,所以尼安德特人应是演化中的一个旁支。这两具头骨显示出有较圆的头颅和较高的颅顶。虽然今天大家都承认这两具头骨上直立人的性状仍很明显,但是在当时却形成了一种叫“人科灾变说”的理论,认为以这两具头骨为代表的晚期智人大约在20万年前已经出现。可是他们受到尼安德特人的压制,因为尼安德特人在体质上更加适应严酷的冰期气候,而体质形态上更为进步的晚期智人只是生存在亚洲一些较为温暖的地区,逐渐发展出更为进步的技术和文化。大约在3.7万年前,他们返回欧洲取代并消灭了尼人。
“非洲起源说”20世纪80年代以后也被称作“夏娃假说”,最早于20世纪70年代中期提出,80年代以后由于对各大洲现代人DNA序列的研究而得以盛行,之后得到许多考古学家、遗传学家及分子生物学家研究结果的证实。支持该学说的研究者认为,现在世界各地的解剖学上的现代人不是由当地的古人类直接进化而来,而是来源于10~20万年前起源于非洲的共同祖先,该祖先其后向其他大洲扩散并先后取代了当地的古人类。通过对最近几年有代表性的几例DNA、Y染色体的SRY及YAP区DNA序列、X连锁基因(Xq13.3、PDHA1)及常染色体控制区基因(Mx1、Ede、Ecp、Mcir和Ms205)的序列构建的系统发育树的统计学分析认为,非洲人对现代人类基因组的贡献是通过至少两次向世界其他地区的领域大规模扩张而完成的。
体质上完全具现代性状的人类确实在7万~8万年前已出现在非洲,他们的扩散是通过基因交流和人口迁移来完成的。
二、现代人的迁徙
(一)现代人的迁徙
1、迁徙的路线。10万年前有一批非洲人离开非洲大陆,现在所有非非洲人都是他们的后裔。10万年前也许是我们祖先最早离开非洲的时间,此后人类陆续离开非洲,大约一万年前,人类大规模迁出非洲大陆。
我们的祖先“走出非洲”的路线究竟怎样,还是个未解之谜。如(图一)所示,传统学说和基因学推测,人类大约在一万年前由于自然灾害被迫大规模迁出非洲,对盐的需求使得人类必须沿海岸线迁徙。当越过红海和阿拉伯半岛来到伊朗高原,人类文明得到了了进一步发展,于是一部分向欧洲进发,另一部分继续沿海岸线迁徙来到印度和缅甸。有一部分沿横断山脉和青藏高原边缘向北来到黄土高原,他们依靠盐碱地和盐矿获得食盐。另一部分则向东来到南海边,并继续向北达到朝鲜半岛和东北亚,甚至越过白另海峡到达美洲。在东南亚,有一群擅长航海的人,他们驾驶独木舟来到澳洲及太平洋上的许多岛屿,还有一小部分到了南美洲。当然,沿途有许多部落留在当地。
2、山海经。《山海经》是先秦古籍,是一部富于神话传说的最古老的地理书。它通过口口相传记述了古代地理、物产、神话、巫术、宗教等,也包括古史、医药、民俗、民族等方面的内容,最后由西汉的刘歆整理汇编而成。除此之外,《山海经》还记载了一些奇怪的事件,对这些事件至今仍然存在较大的争论。
该书按照地区不按时间把这些事物一一记录。所记事物大部分由南开始,然后向西,再向北,最后到达大陆(九州)中部。九州四周被东海、西海、南海、北海所包围。古代中国也一直把《山海经》当作历史书看待,是中国各代史家的必备参考书,该书成书年代久远,具体时间已无从考证。书中描述的很多事物在中国境内没有,也无法解释,但在世界其他地方却可以解释。
《山海经》中有两段对黑人的描述:雨师妾在其北。其为人黑,两手各操一蛇,左耳有青蛇,右耳有赤蛇。劳民国在其北,其为人黑。在中国境内没有黑人,即使在印度也只有褐色人种。很显然雨师妾和劳民国应该在非洲大陆。
另外,《山海经》中记载有小人国,名靖人。当今世界只有非洲中部丛林和亚洲的安达曼岛有小人部落存在,他们就是俾格米人。
这里有两个疑问,第一是人类源于非洲应该都是黑人,只是生活环境改变,肤色才变浅。第二是小人国可能是中国的古人游历时发现的。
复旦大学生命科学院最新理论:大约15万年前,在东非分化出了很多人种与部落,其中就已经包含了现在的黑、棕、黄、白四个人种的祖先。黄种人走出非洲的时间要比棕色人晚了近5万年。但他们的扩散速度很快。他们只用了棕色人1/5的时间就到达东南亚。
显然,黑人部落在人类迁徙的路线中是没有的。而亚洲的俾格米人同古印度人一样是棕色人种,他们比其他人种更早迁出非洲,在我们祖先向东迁徙时被发现。中国的古人要想在游历时发现小人国,再全身而退,回到中国,记录所见所闻,在当时的条件下根本不可能。
(二)现代人迁徙的原因
1、年轻的尼罗河。尼罗河位于非洲东北部,是一条国际河流。发源于赤道南部东非高原上的布隆迪高地,干流流经布隆迪、卢旺达、坦桑尼亚、乌干达、苏丹和埃及等国,最后注入地中海。支流还流经肯尼亚、埃塞俄比亚和刚果(金)、厄立特里亚等国的部分地区。干流自卡盖拉河源头至入海口,全长6670公里,是世界流程最长的河流。流域面积约287万平方公里,占非洲大陆面积的九分之一以上。入海口处年平均径流量810亿立方米。尼罗河是由卡盖拉河、白尼罗河、青尼罗河三条河流汇流而成。尼罗河下游谷地三角洲则是人类文明的最早发源地之一,古埃及诞生在此。至今,埃及仍有96%的人口和绝大部分工农业生产集中在这里。因此,尼罗河被视为埃及的生命线。几千年来,尼罗河每年6~10月定期泛滥。8月份河水上涨最高时,淹没了河岸两旁的大片田野,之后人们纷纷迁往高处暂住。十月以后,洪水消退,带来了尼罗河肥沃的土壤。在这些肥沃的土壤上,人们栽培了棉花、小麦、水稻和椰枣等农作物。在干旱的沙漠地区上形成了一条“绿色走廊”。而五千年的文明古国——古埃及,就在这里创造出辉煌的埃及文化。现今,埃及90%以上的人口均分布在尼罗河沿岸平原和三角洲地区。埃及人称尼罗河是他们的生命之母。
尼罗河是一条古老的河流,6500万年前的始新世就已存在。但是尼罗河又是一条年轻的河流,现在的尼罗河大约在1万~1.2万年前出现一次特大规模的泛滥,河流改道,水域生态发生较大变化。这次泛滥持续时间较长,也许有几百年甚至一两千年。大泛滥后,人类开始迁徙,不久水稻和麦子等农作物出现。
2、大洪水的传说。洪水神话在世界上许多国家和民族中几乎普遍存在。在中国有十几个少数民族有一个共同的“葫芦神话”。神话讲述的是远古时期,发生了特大的洪水灾害,人类几乎灭绝,唯有一对兄妹躲在葫芦中得以逃生,洪水退却,兄妹二人结为夫妻,人类获得繁衍。而汉族几乎有一个相同的神话,即伏羲和女娲兄妹成婚的故事。
而在西方国家,洪水神话同样存在,挪亚方舟就是一个典型的故事。挪亚是个义人,在当时是个完美的人。那个时代的世界充满了罪恶,上帝后悔造人,决心制造洪水使他所创造的一切生命都被毁灭。上帝看挪亚是个义人,就想保他全家性命。提前让挪亚造一个方舟。等方舟造好后,果然天降洪水。挪亚就带着他的家人以及牲畜躲进了方舟。洪水泛滥在地上四十天,水往上涨,方舟在水面上漂来漂去。水势在地上极其浩大,天下的高山都淹没了。水势比山高过十五肘,山岭都淹没了。凡在地上的所有生命都死了。只有进方舟的挪亚一家还活着。
世界各大民族的洪水传说并不是说居住在世界各地的人类同时遭遇大洪水,除非整的地球被水淹没。很明显,现代人起源于非洲大陆,尼罗河的泛滥迫使人类大规模迁往世界各地,同时也把文明带到世界各地。
三、北京猿人是不是中国人的祖先
(一)现代人起源的争议
50万年前的北京猿人是不是中国人的祖先?学术界一直存在着很大的争议。争论的一方为古人类学家,他们认为,约50万年前的北京猿人是中国人的祖先。而争论的另一方为遗传生物学家,他们认为,北京猿人并不是我们的祖先,中国人最早的祖先是大约6万年前进入东亚的非洲人。我们姑且把古人类学家的观点称为“多地区起源论”,即现代人是多个地区的古猿独立演化的。而遗传学家的观点就是“单一地区起源论”,即现代人是一个祖先演化而来的,这里单指“非洲起源说”。
1、多地区起源论。这个理论的主要观点是,在100万~200万年前,直立人由非洲扩展到其他大陆后,分别独立演化为现代非洲、亚洲、大洋洲、欧洲人。也有观点认为,与非洲一样,亚洲、欧洲,甚至大洋州都是人类起源的中心,它们完全是按照自己的历史来演化的。而在中国最主要的证据是,中国目前发现的古人类化石,有200多万年前的巫山人、湖北建始人、170多万年前的元谋人、115万年前的蓝田人、50万年前的北京人、35万年前的南京人、30万年前的和县人、十几万年前的长阳智人、2万多年前山顶洞人等古人类遗迹。时间跨度从200万年到1万多年的化石证据都没有间断过,从原始人类到现代人类的演化进展是连续的,这在一定程度上证明,中国人是自己演化而来的,与“非洲人”无关。
2、单一地区起源论。较早提出现代人“非洲起源说”的是美国的两位科学家华莱士和威尔逊,他们在1987年分别带领两个实验室通过检测细胞线粒体内的遗传物质脱氧核糖核酸发现,现代人祖先可追溯到大约15万年前非洲的一个女人“夏娃”。“夏娃”的后裔开始由非洲大陆向世界其他各洲迁移。至于其他各洲的原始人,有一些科学家推断他们被冰川严寒全部自然消灭,也有一些科学家推断他们全被夏娃的后裔征服并取代。关于夏娃的后裔最早来到中国的时间,大约是在5~6万年前,他们来到中国定居下来,生息繁衍,并取代了原来生活在中国大陆的原始人。
在2005年3月7日《科学》杂志上公布的文章,剑桥大学的三位研究人员对现代人类从非洲大陆向世界其他各洲迁移的地理位置和相应的人口基因进行分析研究。结果发现距离埃塞俄比亚(非洲东部国家)的人与埃塞俄比亚人间呈现出很小基因变异性。现代人类开始由非洲大陆向世界其他各洲迁移的路线上,基因多样性的特征表现不明显,没有出现大量的基因中断。这成为进一步证明现代人“非洲起源说”的证据。
(二)基因研究得出的结论
1、基因研究的原理。基因工程科学,是考古研究的重要手段。用DNA可以解决人种起源和繁衍分支的许多问题。生物的遗传基因很可能发生突变或者错误复制,这些情况会原封不动传递给后代,留下了鲜明的排列口令记录。人类基因排列的多样性,就是因为基因复制过程中出现了突变或者错误所造成的,每一个人体的基因组共有三十亿个核酸,发生传递错误或然率为千分之一,故多态性位点达三百万个,分析DNA的多态性,可以向上追溯人类的祖先。人类有二十三对染色体,其中二十二对可以配对,所以出现了DNA交换,这就是“重组”。存在细胞质内的环状DNA,就是线粒体DNA,研究线粒体的进化,就可以知道人类所有成员的母亲到底是谁?男性体内以有Y染色体,它不会发生DNA重组,Y染色体比线粒体长很多,但是分析男性DNA顺序的工作也要复杂得多,只有在DNA测序手段成本便宜的今天,才可以进行这种染色体分析。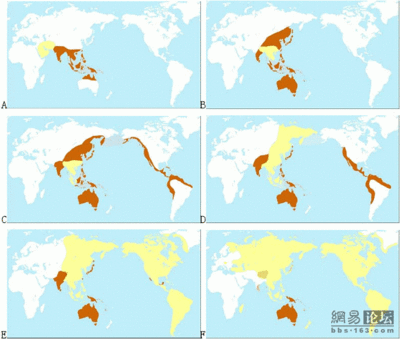
2、尼安德特人。经过分析,所有人类的母系都来自非洲,所有人类的父系都来自非洲。经过对于在克罗地亚发现的尼安德特人头骨的分析,早在二百万年前,非洲猿人已经进化为直立人,并且开始向全世界扩散。在一百万年前,一支直立人进入欧洲,适应了欧洲严寒的天气,进化为身材较矮(约五尺三寸)、但非常粗壮的尼安德特人,另一支迁徙至北京在周口店定居,成为著名的北京人。两者是兄弟,都不能进化为现代智人。随着DNA技术的进步,追宗认祖的亲子鉴定已可揭开人类祖先的真正身份。
现代人体内没有尼安德特人的线粒体DNA。尼安德特人没有对现代人的基因造成任何影响。尼安德特人和克罗马农人共存了很长一段时间,两个不同的人种在一起,矛盾是不可避免的,但融合也肯定是不可避免的。为什么尼安德特人没对现代人的基因造成任何影响?是否说明尼安德特人和克罗马农人根本不能结合产下有生育能力的后代。虽然曾经发现过头部有尼安德特人特征,身体有克罗马农人特征的混血后代,但这个混血儿应该是没有繁育能力的。尼安德特人和克罗马农人与马和驴一样,能产下后代,但它们的后代骡子是没有生殖能力的。
如果人类是在各地起源的,那他们的基因差异至少在百万年,他们应该早已因为生殖隔离(就如同马和驴)而不能互相通婚产生混血后裔。如果已经灭绝的尼安德特人还不够说服力,那我们谈谈现在的热门话题混血猫科动物,且不说分化达200万年的虎和狮,就算亲缘关系最近的豹和狮,它们是在120万年前由同一种猫科动物分别演化而成的,但它们的后代是没有完全生育能力的,也就是存在生殖隔离印度和日本的动物园都曾培育过豹狮兽。一般而言,变种间的杂交是能育的,它们的杂种后代也能育,而物种间的杂交是不育的,物种和杂交种之间可划出一个明确而清楚的界限。
3、北京猿人不是中国人的祖先。同样的道理,50万年前的北京猿人作为洞穴人与其他猿人没有基因交流,已经是一个独立的物种,而一个中国人如果娶一个欧洲女子为妻,则毫无疑问的可以正常生儿育女,繁衍后代,这也说明现代人不管是什么肤色,都属于同一物种。所以北京猿人并不是蒙古人种的祖先。北京猿人虽然一百多万年前已经来到华北平原,但是没有演化成为人类。四万年前,北京猿人已经灭绝了。取而代之的是从非洲迁徙来到亚洲的智人,他们的大脑进化、智力水平、说话能力、群体组织、创造工具能力和战争能力都比北京猿人(直立人)更加优秀,优胜劣汰,北京直立人退出了历史舞台,只是在周口店洞穴中留下了大名鼎鼎的头盖骨。真正的中国人来自于非洲,龙和狮的传说就是最好的证明。世界上没有龙,只有中国古代的传说中有。但是如果见过印度的长吻鳄就会发现两者惊人的相似,只是长吻鳄头上没有角,龙的原型毫无疑问就是鳄鱼。而中国的狮子也只是传说,中国并不出产狮子,而狮子只有非洲和印度才有。或许年代久远,又是口口相传,中国的石狮子和真狮子相貌才差别较大。
四、中华民族的形成
(一)起源于缅甸
1、基因研究的证明。复旦大学的研究显示含M122突变的群体中得出M122C突变发生的年代大致是在18,000~60,000年前,在汉族和藏缅民族的M122突变个体中计算得出两者的分化时间为5,000年(此年代由DYS390推算而得,是三个微卫星位点年代中最大的)。一般而言,单纯从遗传学角度对古代人类的群体迁移和基因突变年代的精确估算困难非常大,因为在上述公式中有效群体大小和突变率的估计带来的偏差直接影响结果,但做出这样的年代估算与体质形态学和考古学在齿型、石器和化石的发现相符。
考古学研究表明涵盖中国北方地区的北亚人种特有的Sinodont齿型发生在1.8万~2.5万年前,而这种齿型是由东南亚地区的类Sinodont齿型演变形成的,这与我们用公式推算出的M122C发生年代下限是相符的。此外,在西伯利亚、贝加尔湖和阿尔泰地区近来的考古学研究表明,这一地区的人类石器文明出现在2.5万~4.5万年前。如果西伯利亚和整个东亚群体由非洲起源并经东南亚迁移而形成,那么现代人类进入东亚地区应该在北亚出现石器文明之前。认为M122C突变估算年代的上限6万年前更接近于事实了,因此研究认为1.8万-6万年是早期现代人迁入东亚所造成的瓶颈效应的年代。据此,该研究认为随着冰川期逐渐消亡,非洲起源的现代人约在6万年前从南方进入东亚,在以后的数万年中逐渐向北迁移,遍及中国大陆,北及西伯利亚。大约在8500年前,非洲来的新移民把文明带到了东亚,以仰韶文化为代表的最早的中华文明开始在黄河中上游地区萌芽。
2、语言学的推测。汉族今天能成为几乎占了全世界人口五分之一的世界第一大民族,这与在历史上优秀的汉文化曾影响了生活在同一环境的其他兄弟民族甚至包括外来的民族,最后他们都接受了汉文化,今天成为中华民族的一员有关。今天汉人的祖源实际上是多来源融合的,但最早说原始汉语的汉祖起着主导作用,他们是来自西南边陲的。汉语和缅甸语同为汉藏语系,语言学家推测汉族起源于缅甸,这也在一定程度上验证了生物遗传学的研究成果。
(二)进入中国
中国利用DNA分析自己祖先的迁徙路线已经相对比较成熟了,中科院云南动物研究所,哈尔滨医科大学,中科院细胞遗传学院等都在进行这方面的研究,只要把各自分散的路线拼合起来就是一条完整的中国人迁徙路线。就目前的研究成果来看,华夏56个民族和东亚、东南亚各民族都是由南亚语系的先民分化出来的,因为他们身上都带有M122的突变。
M122突变大约发生在3万年前,那时中国陆地上的许多山脉被积雪常年覆盖。除了有少数棕色人在黄河、长江流域活动,更多的地方显得空茫一片。这时候,在东南亚的缅甸一带,黄种人的部落已经取得一定势力,棕色人不断向南退缩。如(图二)所示当中国陆地上的冰川不断地消融时,一支带有M122突变的南亚语人群开始进入了中国。目前发现关于南亚语先民进入中国后的分化路线有3条,共有2个入口,一个是在云南,一个是珠江流域。
其中一支沿着云贵高原西侧向北跋涉,他们最终在距今1万年前的时候到达了河套地区,黄河中上游的盆地。这批汉藏语系的祖先也被后人称之为先羌,他们也就是汉族与藏族人的共同祖先。这一支南亚先民在出发时的头颅还很圆,又因为地中海贫血基因的关系,出现了大鼻子、厚嘴唇等特征。但他们顺着江,走着河谷,从云南来到甘肃,人体形态也发生变化。而到了高原之后,不再受疟疾影响,加上缺氧,地中海贫血基因的人不容易存活。于是存活下来的人,脸都变得很长,线条、棱角变得刚硬、如同刀削。
在漫长的行进中,他们使用的语言也发生变化。有一种理论认为,这是口耳相传所带来的变化。听的人觉得你在讲另一个字,复述的时候就变成另外一个音了。更重要的是,经过这1万多年的迁徙,在他们的体内M122的基础上又诞生了一个新的突变M134。这样一直到5000到6000年前,人体内部的DNA又开始“不安分”了,那时,由于粟谷农业的出现,新石器文化开始在这个地区发展。人口的增长使群体必须扩增新的居住地。汉藏语系的两个语族开始分野。其中一个亚群在M134的基础上又发生了M117的突变。他们带着这个突变向东行走,一直到渭河流域才停留下来。他们掌握了农业文明,开始以农耕为生。这个群体就是华人,也就是后来所称的汉人。M117是汉人身上很古老的一个遗传突变。
当南亚先民沿云贵高原西侧北上形成汉藏语系的同时,从中分离出一支南亚先民朝东南方向迁徙,绕过老挝的崇山峻岭,距今1万多年前,南方的北部湾形成了澳泰语系祖先。这一支黄种人带着M119突变从越南、广西方向进入中国,沿着海岸线往东北走,形成了百越民族。具有M119突变的族群在民族学上又称为百越民族系统,他们历史上的覆盖范围是从越南北部的交趾分布到江浙一带。在千百年的分化中,诞生了黎族、侗族、水族、仫佬族、仡佬族、高山族、壮族、傣族。这些民族在语言上、文化上有很强的一致性,所以也统称越人。
有学者在上海调查吴人和越人究竟是一个民族还是完全分属两个民族。之前在语言学界有观点认为他们同属一个民族,因为他们之间的语言能相互沟通。经研究发现,他们完全是两个不同的民族。越人在七八千年前就在上海的松江发展。吴人要到3000年前才进入上海。越人各民族的身上有大量的M119突变。吴人则很少有,他们更多的是M7突变,那是从苗瑶语系分化出来的。当初南亚先民从云贵高贵西侧进入中国后,还有一支人群与汉藏民族分开了,他们沿着长江往下走,这个时候,他们DNA链上位点在M134的基础上发生了M7的突变。这个人群在洞庭一带形成了苗瑶语系。吴人就是苗瑶人东进和南下的汉人同化后形成的,所以他们遗传结构的变化多而复杂。
新的研究发现,在2万多年前,澳泰语系形成的时候,就有一支部落沿着海岸线往上跑。几乎就没有留下沿途停滞的痕迹。他们一直跑到西辽河流域再停留下来。他们是“阿尔泰”语系的核心。后来夷人上去了,华人也上去了,在那里进行了一次民族大融合。形成兴隆文化。这是目前发现的中国最早进入新石器文化的地方。之后那些“阿尔泰”语系的先民又朝周边迁徙,往西分化成蒙古和突厥,往东进入朝鲜和日本,向北穿过白令海峡踏上美洲。
(三)炎帝、蚩尤和黄帝
1、炎帝。炎帝,传说上古时期姜姓部落的首领,又称赤帝、烈山氏,一说即神农氏(或神农氏的子孙)。相传其母名任姒,一日游华山,看见一条神龙,身体马上有反应,回来就生下炎帝。炎帝生于烈山石室,长于姜水,有圣德,以火德王,故号炎帝。炎帝少而聪颖,三天能说话,五天能走路,三年知稼穑之事。他一生为百姓办了许多好事:教百姓耕作,百姓得以丰食足衣;为了让百姓不受病疾之苦,他尝遍了各种药材,以致自己一日中七十次毒。他又作乐器,让百姓懂得礼仪,为后世所称道。
2、蚩尤。蚩尤是中国苗族的先祖。因为现在苗族的族属渊源和远古时代的“九黎、三苗、南蛮”有着密切的一脉相承的关系。在原始社会末期,在我国长江中下游和黄河下游一带,远古的时候就生活着很多原始人类;他们经过世世代代的生息繁衍,通过艰苦的劳动,在距今五千多年前,逐渐形成了部落联盟。这个部落联盟叫做“九黎”,以蚩尤为首领。《国语•梦语》注中说:“九黎,蚩尤之徒也”。《书吕刑释文》、《吕氏春秋•荡兵》和《战国策•秦》中,都说蚩尤是九黎之君。他们借助优越的地理条件,不断地辛勤开拓,使生产力不断提高,社会经济不断发展,一跃而成为雄踞东方的强大部落,并且最早进入中原。著名历史学家范文澜同志写道:传说中的中国远古居民,“居住在南方的人统被称为‘蛮族’。其中九黎族最早进入中部地区。九黎当是九个部落的联盟,每个部落又包含九个兄弟氏族,共八十一个兄弟氏族。蚩尤是九黎族的首领,兄弟八十一人,即八十一个氏族酋长。……是以猛兽为图腾,勇悍善斗的强大部落。”蚩尤发明了金属冶炼和金属兵器的制造。所以在蚩尤死后,黄帝及其后代帝王都把蚩尤奉为“兵主”,视为“战神”来崇敬和缅怀。
3、黄帝。黄帝与炎帝都被看作是华夏民族的始祖,故中国人有时自称“炎黄子孙”。传说中黄帝的正妃是嫘祖,次妃为方雷氏,彤鱼氏和嫫母。据《史记》记载:“黄帝二十五子,得其姓者十四人。”颛顼、帝喾、唐尧、虞舜,以及夏朝、商朝、周朝的君主都是黄帝的子孙。
关于黄帝的传说:据传他出生几十天就会说话,少年时思维敏捷,青年时敦厚能干,成年后聪明坚毅。建国于有熊(河南新郑),亦称为有熊氏。黄帝在位时间很久,国势强盛,政治安定,文化进步,有许多发明和制作,如文字、音乐、历数、宫室、舟车、衣裳和指南车等。相传尧、舜、禹、汤等均是他的后裔,因此黄帝被奉为中华民族的共同始祖和人文祖先。
4、涿鹿之战。传说距今约4600余年前黄帝联合炎帝部族与东夷集团九黎族首领蚩尤在涿鹿(今河北省涿鹿县)进行的一次艰苦激烈的大战,这是远古时代一次很大规模的战争。相传神农氏为帝王时,各氏族、部落相互争战。阪泉之战后,黄帝势力大增,蚩尤不肯听命。黄帝征集各部落、联盟兵众,与蚩尤大战于涿鹿(今河北涿鹿东南)之野。相传黄帝命应龙作水阵阻挡蚩尤,但被效忠于蚩尤的风伯雨师所破。黄帝又请旱神女魃作法,天气骤然放晴,旱热难当,使久习于东南方阴雨气候的蚩尤无法忍受,败阵南逃,于冀州之野擒杀蚩尤。“战争”的目的,是双方为了争夺适于牧放和浅耕的中原地带。它也是我国历史上见于记载的最早的“战争”,对于古代华夏族由野蛮时代向文明时代的转变产生过重大的影响。九黎族原居于南方,后北上中原,与以黄帝为首的部落发生了冲突,双方战于涿鹿之野。据说蚩尤族善于制作兵器,其铜制兵器精良坚利,且部众勇猛剽悍,生性善战,擅长角牴,进入华北地区后,首先与炎帝部族发生了正面冲突。蚩尤族联合巨人夸父部族和三苗一部,用武力击败了炎帝族,并进而占据了炎帝族居住的“九隅”,即“九州”。
炎帝族为了维持生存,遂向同集团的黄帝族求援。黄帝族为了维护华夏集团的整体利益,就答应炎帝族的请求,将势力推向东方。这样,便同正乘势向西北推进的蚩尤族在涿鹿地区相遭遇了。当时蚩尤族集结了所属的81个支族(一说72族),在力量上占据某种优势,所以,双方接触后,蚩尤族便倚仗人多势众、武器优良等条件,主动向黄帝族发起攻击。黄帝族则率领以熊、罴、狼、豹、雕、龙、鸮等为图腾的氏族,迎战蚩尤族,并让“应龙高水”,即利用位处上流的条件,在河流上筑土坝蓄水,以阻挡蚩尤族的进攻。
战争爆发后,适逢浓雾和大风暴雨天气,这很适合来自东方多雨环境的蚩尤族展开军事行动。所以在初战阶段,适合于晴天气环境作战的黄帝族处境并不有利,曾经九战而九败(九是虚数,形容次数之多)。然而,不多久,雨季过去,天气放晴,这就给黄帝族转败为胜提供了重要契机。黄帝族把握战机,在玄女族的支援下,乘势向蚩尤族发动反击。其利用特殊有利的天候——狂风大作,尘沙漫天,吹号角,击战鼓,乘蚩尤族部众迷乱、震慑之际,以指南车指示方向,驱众向蚩尤族进攻,终于一举击败敌人,并在冀州之野(即冀州,今河北地区)擒杀其首领蚩尤。涿鹿之战就这样以黄帝族的胜利而宣告结束。
这场“战争”的大致经过情况是由神话传说所透露的,因此更具体的细节已无从考索了。但是神话毕竟是历史的投影,曲折地反映了事实的本身。从这个意义上说,涿鹿之战堪称为我国古代战争的滥觞。涿鹿之战中,黄帝族之所以取得最后胜利,在于其战争指导比蚩尤族要来得高明。具体而言,即其已开始注意从政治和军事两方面作好战争准备,史称“轩辕氏乃修德振兵”,就是证明。在战争过程中,黄帝族还善于争取同盟者,并能注意选择和准备战场,巧妙利用有利于己不利于敌的天候条件,果断及时进行反击,从而一举击败强劲的对手,建立自己对中原地区的控制。而蚩尤族虽然人数众多,武器先进,但可能处于军事转型期,即石制兵器向金属兵器的转变,由于金属兵器制造工艺还不成熟,成本较高,不能象石制兵器那样就地补给,所以在这场空前规模的战争中败下阵来。涿鹿之战,有力地奠定了华夏集团据有广大中原地区的基础,并起到了进一步融合各氏族部落的催化作用。取得这场战争胜利的部族首领黄帝从此成为中华民族的共同祖先,并被逐步神化。由此可见,涿鹿之战的确为我们中华民族在发轫时期决定日后基本面貌的历史性“战争”。
(四)中国历史上的游牧民族
1、消失匈奴。中国部分史籍记载,匈奴人是夏朝的遗民。《史记•匈奴列传》记载∶“匈奴,其先祖夏后氏之苗裔也,曰淳维。”。《山海经•大荒北经》称∶犬戎与夏人同祖,皆出于黄帝。《史记索隐》引张晏的话说∶“淳维以殷时奔北边。”意即夏的后裔淳维,在商朝时逃到北边,子孙繁衍成了匈奴。还有一说认为,移居北地的夏之后裔,是夏桀的儿子。夏桀流放三年而死,其子獯鬻带著父亲留下的妻妾,避居北野,随畜移徙,即是中国所称的匈奴。部分学者根据《史记》记载的後半段文字,认为匈奴原是山戎、猃狁、荤粥。王国维在《鬼方昆夷猃狁考》中,把匈奴名称的演变作了系统的概括,认为商朝时的鬼方、混夷、獯鬻,周朝时的猃狁,春秋时的戎、狄,战国时的胡,都是后世所谓的匈奴。还有一说,把鬼戎、义渠、燕京、余无、楼烦、大荔等史籍中所见之异民族,统称为匈奴。还有人认为匈奴与先秦时期的北方少数民族不可混为一谈,匈奴应是西方草原的一个游牧民族,战国末期之前,还未游牧至中国北部。战国末,常扰掠秦、赵、燕北边,三国相继筑长城以拒之。冒顿单于在位(前209~前174)时,统一各部,建立国家,统有大漠南北广大地区。老上单于(约前174~前160)时,匈奴势力东至辽河,西越葱岭,北抵贝加尔湖,南达长城,成为历史上第一个草原游牧帝国。汉初,匈奴不断南下侵掠。公元前200年,围汉高祖刘邦于白登山(今山西大同东北),遂迫汉朝实行和亲,且岁奉贡献,并开关市与之交易。然而,匈奴仍屡屡背约南侵,成为汉朝一大边患。汉武帝时国力强盛,曾3次(前127、前121、前119)大举出兵反击匈奴,匈奴势力渐衰。汉代,匈奴由于天灾、人祸及汉军的打击,发生过两次分裂:一次是公元前57年左右出现的五单于并立局面。结果是公元前53年呼韩邪单于归汉,引众南徙阴山附近。公元前36年,汉西域副校尉陈汤发西域各国兵远征康居,击杀与汉为敌的郅支单于,消灭了匈奴在西域的势力,公元前33年,汉元帝以宫人王嫱(昭君)嫁呼韩邪单于,
公元91年,汉军出居延塞(今内蒙古西部额济纳旗一带),围北匈奴单于于金微山(今阿尔泰山)。北匈奴战败后部分西迁,余众后来归附于兴起的鲜卑。多数学者认为,西迁的北匈奴就是欧洲史上的匈人。南匈奴南下汉化,一直居住在河套一带,三国时期曹操把匈奴分成五个部。4世纪初,匈奴族的五部大都督刘渊在成都王司马颖手下为将。乘西晋八王之乱之后的混乱时期,刘渊起兵占领了北中国的大部分地区,自称汉王,311年攻占洛阳,316年攻占长安,灭西晋。史称前赵或汉赵。
匈奴有一支地位低下的白种人族群称为羯人。汉赵的大将羯人石勒自立,建立赵国,史称石赵或后赵,351年被汉族人冉闵所灭。
融入匈奴人中的月氏人,称为匈奴别部卢水胡。其中沮渠家族推后凉汉官段业为主,在现甘肃地区建立北凉。后沮渠蒙逊杀段业,自立为北凉主。后被鲜卑人拓跋氏北魏所灭。
匈奴与鲜卑的混血后代称为铁弗人。铁弗人刘勃勃被鲜卑拓跋氏击败后投奔羌人的后秦。后自认为是末代的匈奴王,改姓赫连,在河套地区创立夏国,史称胡夏。后被北魏所灭。
匈奴融入靠近高丽的鲜卑的宇文氏部落,进入朝鲜半岛。后来宇文氏篡西魏建立的北周政权,后被汉族外戚杨坚所篡。杨坚创立隋朝,统一中原。
以上是五胡十六国及南北朝时期,匈奴在中国历史舞台上进行了最后一场演出。之后,亚洲部分的匈奴最后一点残余部族,作为一个独立民族的身份从中国历史中彻底宣告终结,和其他一些民族一起消融于汉族为主体的华夏族。
匈奴后裔汉化后,所改汉姓有刘、贺等等,据说有一些目前还生活在今天的陕西、山西等地。
2、汉藏同源。藏族的起源没有特别可靠的文献记载。如果考察风俗习惯的联系,由于各民族间的广泛交流,不会太准。考察DNA的情况也是如此。唯一公认的是藏语与汉语同属“汉藏语系”。这并不常见,象满蒙两个民族与汉族的血统和文化交流要密切得多,但满语蒙语却属于“阿尔泰语系”。
古人认为:藏即羌也。现代中国专家经考证也认可了这一结论。而羌族又是所谓华夏民族的组成部分之一,传说中的大禹就是羌族人。这就形成了极密切的关系。DNA方面的结果说明了藏族是“蒙古人种”,
无论是从外貌到文化,都很难让人相信汉人和藏人本出同源,但分子人类学家对此深信不疑。最早提出汉藏同源的是中国的一些语言学家。原本在语言学界认为南方的侗傣语和汉语最接近,后来发现与汉语最接近的是藏语。
藏语中大多数字的发音在中国东南部的方言中都能找到,而且字义相同。藏语和汉语之间存在的不是一般意义的对应关系,而是系统性的一一对应。在语言上,汉族和藏族表露出非同一般的关系。
中科院昆明动物研究所于1996年在DNA中找到了汉藏同源的证据。发现汉人和藏人在M122以及在其分支M134上都有相同的突变。通过对Y染色体主成分分析,发现藏族的突变频率与汉人最接近,这意味着在藏缅语系的诸多民族中,藏族和汉族最接近。他们分化的年代约在5000年前。
3、幸存的蒙古。蒙古族是有悠久历史和灿烂文化的民族,传说中的蒙古人已有三千多年的历史;据文字记载,也有一千多年的历史了。
蒙古族的族源问题,现在多数的学者都认为:蒙古族属东胡族系。公元前209年,东胡被匈奴冒顿单于所破,东胡诸部在匈奴人统治下达三世纪之久,公元一世纪末至二世纪初,匈奴为汉朝所破,东胡人的一支鲜卑人自潢水流域转徙其地,剩余的匈奴人也都自称为鲜卑,鲜卑自此强盛起来。鲜卑起源自黄帝之孙曰始均,《晋书》记载:曾在晋代受封为“鲜卑都督”的慕容廆,“昌黎棘城鲜卑人也。其先有熊氏之苗裔,世居北夷,号曰东胡。”有熊氏,就是黄帝部落。公元四世纪中叶,居住在潢水、老哈河流域一带的鲜卑人的一支,自称为“契丹”;居住于兴安岭以西(今呼伦贝尔地区)的鲜卑人的一支则称为“室韦”。蒙古部就是室韦人的一支,在唐朝时已有记载,称为“蒙兀室韦”。
据《史集》记载,蒙古部最初只包括捏古斯和乞颜两个氏族,他们被突厥部落打败后只剩下两男两女,他们逃到了额尔古涅昆(额尔古纳河畔山岭)一带居住下来,生息繁衍,大约经过了400多年的时间,部落逐渐兴盛起来,并产生了许多分支,公元8世纪,由于人口的不断增长,为了更好的发展,不得不外迁徙,这时已分出了70个分支了,这70个分支被称为“迭儿勒勤蒙古”。在迁出的蒙古人当中,有一位很有声望的人,名叫孛儿帖赤那,以他为首的迭儿勒勤蒙古自称为“乞牙惕氏”(乞颜的复数)。“乞牙惕氏”人迁徙到了斡难河源头肯特山一带,
据《蒙古秘史》记载,孛儿帖赤那的的十二世孙朵奔篾儿干死后,他的寡妻阿阑豁阿又生了三个儿子,传说这三个儿子是感光而生的“天子”,因为他们是从阿阑豁阿洁白的腰里出生的,因此他们的后裔被称为“尼伦蒙古”。在尼伦蒙古中,以孛端察儿为始祖的孛儿只斤氏就是成吉思汗的祖先。迭儿勒勤蒙古和尼伦蒙古,被统称为“也克蒙古”(大蒙古),他们是原蒙古人。十三世纪初,成吉思汗统一蒙古高原诸部,建立大蒙古帝国。用新的千户制体系分封人口。战败的部落,如塔塔儿、克烈、乃蛮被瓜分到各千户。族属不同、社会发展不平衡、方言各异的各部在统一的汗权统治下,形成了具有共同地域、共同经济基础、共同语言和共同心理素质的民族共同体——蒙古族。
五、结束语
历史上,喜欢把别人归类的习惯给人类带来了不少的苦难。整个族群被屠戮或被奴役,原因只在他们的肤色或眼睛的形状。科学证实所有人类都彼此相连,无论是强盛不衰的中国人、在各地殖民的欧洲人、散布全球的犹太人,还是猎捕羚羊的布须曼人,全都是人类大家庭的成员。在中国,无论是哪个民族,都有着共同的祖先。外貌、语言、文化和习俗的差异都不足以分裂我们的中华民族。展望未来,在科学技术高速发展之下,中华民族的联系将更加紧密,人类大同的新世界已亦将指日可待。
《李辉:黄色人种的起源与扩散》
一、华夏人类的起源【体质人类学基本知识】在本文的一开始,首先需要纠正大家的一个传统观念:中国人皮肤的颜色不是黄色的,而是浅棕色的。中国人眼睛的颜色不是黑色的,而是深棕色的。中国人的头发的颜色也不是纯黑色的,而是带有一点深棕色的。对于中国人种族上的这种性状特征,我们需要了解它的来源。在此之前,我们又必须了解中国人的种族构成。中国人的种族来源,由两大部分构成,一部分是欧亚部落的血统,一部分是古亚洲人的血统。古亚洲人是第一次出非洲的人群,而欧亚人是第二次出非洲的人群中的第二次扩张的人群。古亚洲人分为两种,一种是矮黑人(D-YAP),他们和非洲黑人(尼格罗人)拥有一个共同的祖先,另一种是棕种人(C-M130)。纯种的矮黑人是印度的安达曼人,东南亚的维达人等,而纯种的棕种人如今已不存在,但他们是蒙古人、女真人、澳大利亚土著等民族的直系祖先。矮黑人、棕种人先后从亚洲南部北上东亚后,肤色变浅。矮黑人中最进化的一支是日本的阿伊努人(虾夷人),它们是绳文人的后裔,也是大和民族的重要底子。另外,西藏人中也拥有大量的矮黑人父系成分。第二次出非洲的人群被称为中东部落(F-M89),其中的一支演化为欧亚部落(K-M9),其余进化成地中海-高加索人种,属于暗白人种。欧亚部落的原始人种属于未分离的黄白人种,又演化成好几支。其中一支形成东亚部落(NO-M214),被称为黄种人(又称华夏-芬兰人种),另有一支形成中亚部落,属于早期的白种人。东亚部落的黄种人又分离为两支,一支是芬兰人,部分北亚人的直系祖先(N-M231)。另一支则形成中国人,东南亚人的直系祖先(O-M175)。在M175的基础上,东亚地区的黄种人演化为华夏、东夷、百越、苗瑶、百濮、南岛等一系列民族。而中亚部落也分化为两支,一支是印第安人和他们的北亚祖先(Q-P36),一支是主流欧洲人和印度雅利安人的祖先(R-M207)。其中进入欧洲的白种人是后来的日耳曼人,斯拉夫人,波罗的人,克尔特人的主要祖先(当然这些民族还有少部分其他来源)。这些生活在欧洲北部的人群是相对比较纯的欧亚部落后裔(主要是中亚部落,少数属于东亚部落),他们被称为诺迪克人种(又称北欧人种,白海-波罗的人种),属于显白人种,也是狭义上的白种人。他们和南欧的地中海人属于不同的人种。在混血程度不高的史前,相比于纯种的地中海人和阿拉伯人,纯种的诺迪克—雅利安人和东亚的纯种黄种人血缘关系要更接近。这一点非常重要,也是下文讨论的基础。北欧的诺迪克人种和南欧的的地中海人种在欧洲中部地区发生混血,形成了阿尔卑斯人种。一般认为,诺迪克人是典型的白种人,而阿尔卑斯人是典型的欧洲人,地中海人则接近中东人。亚洲黄种人和亚洲棕种人、矮黑人大约在10万年前就已经分离;亚洲黄种人和北欧白种人大约在4万年前的伊朗高原上分离,而传统认为东亚的黄种人和欧亚大陆北部的黄种人大约在3.5万年在阿富汗分离。不过最新的研究显示这两者的实际分离时间可能比原来的认识大大缩短,并且分离地点很可能在云南一带。另外需要说明,中国人也有部分芬兰-乌拉尔黄种人的血统,不过和欧洲黄种人属于不同的子类型。这些也非常的重要。回到中国,我们简要得概括一下中国人大体上的血缘构成:父系=压倒性的黄种男人(F系下的K系)+极少量矮黑男人(M168下的C)+极少量棕种男人(M168下的D)+极少量的暗白种男人(M168下的F系)+极少量的显白种男人(F系下的K系)((黄种成分南北差异不大))
母系=部分黄种女人(N系下的R系)+部分矮黑女人(非非洲人的M系)+部分棕种女人(非非洲人的M系)+少量暗白种女人(非非洲人的N系)+可能的极少量的显白种女人(N系下的R系)((黄种成分由南向北递减))
以上是以父系的Y染色体遗传标记(父传子不传女,子传孙)和母系的mtDNA遗传标记(母传子女,女传后代,但子不能传后代)为标准的大致比例构成。但决定体质的主要是常染色体。常染色体是一个人父亲家族的所有直系祖先和母亲家族的所有直系祖先的混合,但比较容易被环境改变而不太稳定。中国人身上具体到底有多少是欧亚人血统,有多少是古亚洲人血统,这是一个很复杂的,至今还未有定论的问题。但可以肯定南方人是比北方人更纯的黄种人,中国最纯的黄种人是在两广一带,这些是没有任何问题的。虽然南北汉族父系有80%以上的同宗,母系也有50%的同宗,但是从各种方法研究常染色体的结果来看,北方汉族和北方少数民族,日韩人棕种成分比较多,并且聚类在一起,南方汉族和东南亚人的黄种成分比较多,并聚类在一起,两者之间以长江为界径渭分明。这说明南方人总体上的的黄种血统普遍要多于棕黑血统,而北方人则不好说(其实这个结论从母系遗传的线粒体DNA各类型的分布情况就可以得出了)。由于历史上北方汉族的大规模南下,使得今天的南方汉人融入了更多的古亚洲血统,和东南亚人区分了开来。当然,气候也能影响常染色体的反映状况,这种聚类也有气候的因素。
在开始分析中国人的相貌特点以前,有必要列举新亚洲人(纯黄种人)、古亚洲人(矮黑棕种人)、诺迪克人(北欧人)、地中海人(包括中东人)之间的一些体质上的性状特征如下:
1.纯黄种人:棕色眼睛 双眼皮 大眼睛 两眼居中 中眼窝 中鼻梁 阔鼻头瓜子脸面部线条柔和深棕发直发长头型中颧骨浅棕皮肤身材中等修长 无罗圈腿 体毛较少脑容量最大
2.古亚洲人:黑色眼睛 单眼皮 小眼睛 两眼靠鼻 浅眼窝 扁鼻梁 塌鼻头驴窄脸面部无立体感纯黑发卷发短头型高颧骨深色皮肤身材矮小粗壮 有罗圈腿 体毛发达脑容量一般
3.诺迪克人:蓝灰眼睛 双眼皮 大眼睛 两眼居中 中眼窝 中鼻梁 圆鼻头 介于14面部线条柔和金黄发直发长头型中颧骨白色皮肤身材高大强壮 无罗圈腿 体毛中等 脑容量较大
4.地中海人:棕褐眼睛 双眼皮 大眼睛 两眼居中 深眼窝 高鼻梁 鹰钩鼻椭圆脸面部立体感强棕褐发卷发长头型高颧骨浅棕皮肤身材中等较壮 无罗圈腿 体毛发达 脑容量一般需要说明的是,人类学中一般不用眼睛,皮肤,头发的颜色作为判定种族的根本依据,因为这些在史前的进化过程中是很容易改变的。即使在进化的自然选择作用大大降低的现代社会,一个民族的高矮胖瘦也是比较容易改变的。所以这些都不是根本性的人种特征。判定种族差异的根本区别主要在于人的骨骼方面,尤其是测定各项头骨数据,因为骨骼尤其头骨比较稳定不容易被环境改变。这样,我们可以发现,除去某些处于边缘化的印第安人在北亚的近亲部落以外(北欧人与地中海人混血而成的阿尔卑斯人是后来形成的),东亚的纯黄种人和北欧印度的纯白种人其实是整个欧亚大陆上血统最接近的人种!这不仅在已经遗传科学上获得了证明,而且在体质观察中也可以被确认。写到这里,积累了上面的这些基础知识,现在就可以着手解决标题所提出问题:既然北欧人的祖先和中国人的祖先血缘关系如此接近,为什么中国人长得和北欧人不一样呢?为什么中国人没有强壮的体格,高鼻深目,金发碧眼,白色的皮肤呢?
我国人的种族来源,由两大部分构成,一部分是欧亚部落的血统,一部分是古亚洲人的血统。古亚洲人是第一次出非洲的人群,而欧亚人是第二次出非洲的人群中的第二次扩张的人群。古亚洲人分为两种,一种是矮黑人(D-YAP),他们和非洲黑人(尼格罗人)拥有一个共同的祖先,另一种是棕种人(C-M130)。纯种的矮黑人是印度的安达曼人,东南亚的维达人等,而纯种的棕种人如今已不存在,但他们是蒙古人,女真人,澳大利亚土著等民族的直系祖先。矮黑人,棕种人先后从亚洲南部北上东亚后,肤色变浅。矮黑人中最进化的一支是日本的阿伊努人(虾夷人),它们是纹绳人的后裔,也是大和民族的重要底子。另外,西藏人中也拥有大量的矮黑人父系成分。第二次出非洲的人群被称为中东部落(F-M89),其中的一支演化为欧亚部落(K-M9),其余进化成地中海-高加索人种,属于暗白人种。欧亚部落的原始人种属于未分离的黄白人种,又演化成好几支。其中一支形成东亚部落(NO-M214),被称为黄种人(又称华夏-芬兰人种),另有一支形成中亚部落(P*),属于早期的白种人。东亚部落的黄种人又分离为两支,一支是芬兰人,部分北亚人的直系祖先(N-M231),另一支则形成我国人,东南亚人的直系祖先(O-M175)。在M175的基础上,东亚地区的黄种人演化为华夏,东夷,百越,苗瑶,百濮,南岛等一系列民族。而中亚部落也分化为两支,一支是印第安人和他们的北亚祖先(Q-P36),一支是主流欧洲人和印度雅利安人的祖先(R-M207)。其中进入欧洲的白种人是后来的日耳曼人,斯拉夫人,波罗的人,克尔特人的主要祖先(当然这些民族还有少部分其他来源)。这些生活在欧洲北部的人群是相对比较纯的欧亚部落后裔(主要是中亚部落,少数属于东亚部落),他们被称为诺迪克人种(又称北欧人种,白海-波罗的人种),属于显白人种,也是狭义上的白种人。他们和南欧的地中海人属于不同的人种,在混血程度不高的史前,相比于纯种的地中海人和阿拉伯人,纯种的诺迪克—雅利安人和东亚的纯种黄种人血缘关系要更接近。这一点非常重要,也是下文讨论的基础。北欧的诺迪克人种和南欧的的地中海人种在欧洲中部地区发生混血,形成了阿尔卑斯人种。一般认为,诺迪克人是典型的白种人,而阿尔卑斯人是典型的欧洲人,地中海人则接近中东人。亚洲黄种人和亚洲棕种人矮黑人大约在10万年前就已经分离,而亚洲黄种人和北欧白种人大约在4万年前的伊朗高原上分离,而传统认为东亚的黄种人和欧亚大陆北部的黄种人大约在3.5万年在阿富汗分离,不过最新的研究显示这两者的实际分离时间可能比原来的认识大大缩短,并且分离地点很可能在云南一带。另外需要说明,我国人也有部分芬兰-乌拉尔黄种人的血统,不过和欧洲黄种人属于不同的子类型。这些也非常的重要。回到中国,我们简要得概括一下中国人大体上的血缘构成:其实,中国人的父系祖先还是比较高大强壮的,只是不如白种人(指北欧人种)高大。在生物学中有两个个规律,一个是海拔越高的地方进化成长的物种就越高大粗壮,反之亦然。还有一个是温带地区形成的人种最高大,热带和寒带形成的人种都比较矮小。在人种形成的初期,受外界的自然环境选择作用是很大的。中国人的祖先出非洲后,曾经在伊朗阿富汗一带的中纬度的高原上进化了很长的时间,然后才通过印度缅甸进入云南的中低纬度的高原,又进化了很长的时间,最后才扩散到中国各地,取代了原来生活在中国境内的古亚洲男人。其中华夏族又在西部甘肃的中纬度的高原上生活了很长的时间。北欧人和印第安人的共同祖先和中国人的祖先在伊朗高原上分离以后,进入了中亚的中高纬度的高原地区进化了很长时间。北欧人印度雅利安人的共同祖先接着和印第安人的祖先分离,并分两批向西进入欧洲或印度。第一批进入欧洲的是西欧的克罗马农人,第二批是近两万年后才进入欧洲的从南俄草原进入的作为征服者的印欧人。
印欧人的另一部分向南进入印度,伊朗,阿富汗,他们就是大名鼎鼎的雅利安人。在两批进入欧洲的北欧人之间,隔着一批在农业扩张时代进入欧洲的中东农民,他们带来了欧洲人中的地中海人种的基因,甚至一部分非洲黑人基因(不过和班图黑人属于不同的子类型)。另需提醒,欧洲的纬度高于东亚。史前的欧洲不仅纬度高,而且经历了寒冷的冰川期,地中海人种的某些体质特征的形成某种程度上就拜这次冰川期所赐,克罗马农人也受到过一点影响。地中海人由于生活在炎热的中东时间更长,所以高鼻深目的特征就更突出。印欧人6000年前征服了欧洲后,由于和被征服者克罗马农人,中东农人混血,加上相同地区的相同地理气候条件下产生的趋同进化,使得所有的欧洲人面目特征都看上去比较相似。
在中世纪,欧洲的气候仍然很寒冷,一直到近代大西洋暖流的形成。北欧人虽然长期生活在北欧寒冷的高纬度平原地区,但是由于他们的种族特征主要形成于温带的高原地区,所以并不矮小。中国人的父系祖先也是在高原上进化出的人种,因此先天的身材并不矮小干瘦,只是纬度没有北欧人高,气候条件也比北欧人暖和一些,所以先天基础没有北欧人高大强壮。饮食习惯和营养条件对人种的形成也非常重要。不过因为和身材矮小瘦弱的古亚洲女人混合,导致了后代的中国人身材,体格被大大拉低。但影响身材高大强壮与否的除了人种的先天基础外,还与人种大致形成后的气候条件,饮食习惯和营养条件息息相关。古代北方中国人长期是世界上最高大的人种之一,不亚于欧洲人。倒是欧洲人历史上有一些时期由于营养不佳,身材比今天矮小很多。
再说一下“高鼻深目”的特征。相比于现代东方人的扁平的五官,纯种的黄种人其实是可以称得上“高鼻深目”的,只是没有中东和南欧的地中海人种那么得夸张。“高鼻深目”本来就是M89的中东部落后代的特征(应该来自于非洲祖先),只是地中海—高加索人种实在是进化得有些夸张了。而M9的欧亚部落后代则要柔和很多,这应该和欧亚部落在较高纬度的地区进化过一段时间有关。不过和大饼脸的古亚洲人相比,纯种的黄种人具有明显的欧亚黄白人种的特征,五官立体感要强很多。今天的亚洲基本已经没有纯种的黄种人了,不过这可以参考一下黄种血统比较多的某些东南亚人,以及介于黄白之间的某些北美印第安人。某些东南亚民族,他们是比南方汉族更纯正的黄种人(当然也是和矮黑人棕种人混血过,只是比例要稍低一些)。而南方汉族,是比北方汉族更纯正的黄种人。汉人中黄种血统最少,矮黑人棕种人成分最多的是土生的西北汉族(新疆汉族属于中原移民,另当别论)。另外,南方的四川地区有些特别,矮黑血统比其他南方人要多,这是因为四川是属于矮黑种的历史上的氐人的老家。韩国人类似于西北汉族。而历史上的各种北方胡人(白胡除外,包括满蒙),日本人,基本上不属于严格意义上的黄种人。即使今天的两广人,也不是纯正的黄种人,只是黄种成分比其他中国人更多。很多人污蔑广东人是“马来人”,殊不知,广东人的某些“马来特征”,恰恰是黄种祖先的原始相貌特征的一些反映。二、走向远东的远古人类【肤色与人种的起源】现代人(ModernHuman)的学名叫做晚期智人(Homo sapienssapiens),其生物学级别被假定为一个亚种(subspecies),与各早期智人的亚种同属于一个物种(species)。可能这种级别的设定完全不正确,但至少晚期智人的级别与早期智人的各分支相并列是相当可信的。所以现代人有相当近的同源性,都可以追溯到晚期智人的发源地——非洲古陆。而后在扩散迁徙过程中形成了各分支,我们姑且称其为地理种,即比亚种更小一级的分类单位。不管哪个地理种,与当地出现过的早期智人亚种都没有直系亲缘关系。现存的现代人地理种有4个:即黑色人种(尼格罗人种Negroid)、棕色人种(澳大利亚人种Australoid)、黄色人种(蒙古人种Mongoloid)和白色人种(高加索人种Caucasoid)。虽然肤色是个非常明显的特征,却不是关键的判别特征,同一地理种内的各群体肤色会相差很大,造成有些黑色人种群体比一些白色人种群体还白得多。例如埃塞俄比亚(Ethiopia)的一些黑人群体体色非常白。我们对人种的判定一直采用一种综合的体征标准,以达到确实的亲缘关系。这种标准包括颅面骨骼的各部分形态、发质发色、虹膜色泽,以及其它一些遗传体征。单一的特征都很难判定人种,因为其变异和交流可以造成不同原因的相同表现。比如肤色可能受高紫外线地区的生活环境选择而造成不同的人种都可以有深色的相同表现,不能以肤色深浅判断关系远近。所以所有体征中,颅面形态相对来说是最好的分种依据,因为其受环境影响比较小,变化范围大,变化以后不易回复祖先群体的形态。一般来说,黑人整个头颅较圆,额部较突;白人较侧扁,面部凹凸不平,眶骨深,鼻根低;黄种人前后较扁,面部较平整,鼻根高;棕色人种颅型略偏圆柱形,面部起伏也较大。另外,从发型上看,黄种人多直发,棕种人多窄波发,黑种人多旋发,白种人多宽波发。这些特征一直被体质人类学界用以分析人种。特别是在东亚的人类学研究中,分析棕色人种和黄色人种的交流历史。体质形态的分类主导地位在近二十年内渐渐被单倍遗传系统所取代。所谓单倍遗传系统是指纯由父母之中的唯一一方遗传延续的遗传分子。包括母系遗传的线粒体和父系遗传的Y染色体非重组区。这些单倍分子不受混血影响,其中的无功能区又自由忠实地记录了群体分离的历史。以此判定人种和次人种,层次结构相当清晰。这些人种的分布在地理大发现时代和殖民洪潮之前是相对隔离的。具体分布大致是这样的:黑种人分布于非洲大陆和撒哈拉(Sahara)沙漠以南地区;白种人分布于欧洲、北非、西亚乃至印度;棕种人分布于澳大利亚大陆、新几内亚(NewGuinea)、美拉尼西亚(Melanesia)、安达曼群岛(AndamanIslands)等地;黄种人分布于其余除南极洲以外的地区,包括北亚、中亚、北欧、东亚、东南亚、太平洋诸岛、新西兰(NewZealand)、印度洋诸岛、马达加斯加(Madagascar)和美洲。但在有些地区,可以明显看到人种间普遍的混合特征,比如中亚和北欧,基本上是黄色和白色人种的混合。这种人种混合的体质观察现象由历史记载也可以得到印证。 人种混合的存在提醒了研究远东地区的人类学家。在远东,特别是东南亚,棕色人种和黄色人种错综而居,有些群体更似乎兼有两方的特征,很难判定。这种格局究竟是不是混血,还是其它机制形成的。这个问题引起了人类学界的长期争议。其中的观点基本上可以分三类:气候带说、进化滞留说和南北混合说。气候带说认为,早期的人类分散到各地,长期生活在固定的气候环境中,经过千万年的适应,渐渐形成了不同的体质。这种体质的差异从热带到寒带形成了一个差异梯度,越是往热带越具有棕色人种的特征,越往寒带越具有黄色人种特征。较近的历史时期内的移民活动又局部地打乱了这种南北梯度。气候带说的观点在遗传证据发现之前似乎能解释大部分现象。除了个别案例,比如寒冷的塔斯马尼亚,棕色人种定居历史也相当久远,却没有所谓黄色特征。后来的遗传发现证实了气候带说的观点实为本末倒置。他们所依据的一些特征的确是由气候造成的,却不是长期适应的结果,而是快速选择的结果。比如宽鼻厚唇的热带特征,实际上是由于热带疟166Journal of National Dr. Sun Yat-sen MemorialHall疾高发,而地中海贫血基因可以抵抗疟疾。在强大的生存压力选择下,只要经过数代人的强化和固定,任何群体中的地中海贫血基因都会极高频地出现。而地中海贫血患者的胎儿在发育期间会因为缺氧而造成宽鼻厚唇等特征。这些特征与群体久远的发生历史毫无关系,根本不能解释黄种人和棕种人形成的机理。进化滞留说与结构主义人类学有关系,支持者主要是一些古人类学家。他们认为远东人类由当地的古人类进化而来,其中棕色人种的特征与古人类更接近。他们发现,十万年以来,远东地区的人类化石“渐渐”由棕色人种特征转化为黄色人种特征。旧石器时代基本上“原始”特征占大部分,到新石器时代,“原始”特征个体的比例越来越少。华夏中原地区到夏商时期还有“原始”特征的类澳大利亚个体存在,到后来渐渐消失。这体现了人类的一个进化过程,一些“原始”特征渐渐在群体中被淘汰。而在澳大利亚和伊利安(Irian),“原始”特征却滞留在现代人群中。令人匪夷所思的是,各个人种的进化速度怎么会差别如此大。而各地黄种人却几乎匀速同向地进化了。这种现象的原因根本无法解释,而基因时代是拒绝这样不能解释的理论的。当然,现在的进化研究已经证实了东亚的古人类和现代人无关系,现代人的差异也不可能是因为进化程度的不同造成的。
南北混合说是早期群体遗传学界的普遍观点。他们在大量的双倍遗传标记研究中发现,南北差异始终是主流。对这种现象成因的最自然解释就是南北的人群有完全不同的来源,黄种人来自北方而棕种人来自南方,渐渐地扩散融合形成现代的格局。最早的看法以为现代人是当地起源的,所以猜想黄种人来自北京猿人,棕种人来自爪哇猿人Javaman)。现代人非洲起源说成为共识以后,又猜想黄种人从北方中亚进入,而棕种人从南方东南亚进入。但是从最合适的单倍遗传标记分析看,东亚人群几乎全部是从南方进入而后向北扩散的,所以南北异源之说也无从说起。这种观点基本上是种形而上学的观点。
从单倍遗传标记的分布看,对远东两个人种的分布格局形成机理,可以再提出一种新假说——先后替代说。棕色人种最早到远东,扩散开来;黄色人种来到远东以后渐渐取代棕色人种,形成黄色人种占据主体地区,棕色人种处于偏远地区的格局。
【远东人群遗传层次结构的发现】自从单倍遗传系统的研究建立起来以后,学界对全世界的各个群体作了深入详细的调查分析,数十万的数据呈现在了我们的面前。从这种全面的数据中探索规律,就更能够避免管窥蠡测断章取义了。我们先来看看Y染色体非重组区的单倍群分布,在全世界的分布具有非常明显的地域特异性。
Y染色体主干单倍群在各地人群中的分布比例非洲的E型,美洲的Q型,欧洲的R型,中东的J型都是非常有特色的。这些特色单倍群说明了该区域的人群的主体起源是一致的。更深入地说,在这一地区某种来源的人群的后代已经完全占据了主导地位,其它成或许曾经是多数,但现在已经被覆盖(注:即过去占优势的人种被后来的另一人种基本上消灭掉)。 在非洲,这表现为E对A和B的覆盖;在欧洲则表现为R 对I的覆盖;而在美洲,基本上只能看到Q,其它类群很难观察到。所以在人类定居历史悠久的大陆,一群人扩张开来,替代另一群人的过程往往会发生。这既体现在现代人种对古代人种的替代上,也体现在现代人种的历史中不同人群的更替。族群发生、发展、兴盛最后衰亡的历史在不断的演绎着。
Y染色体主干单倍群的区块分布示意图
在远东地区,这种更替的迹象更为明显。我们可以看到在普遍存在的C和D中间,O型和P型硬生生地插了进来。这种更替的过程并没有完成,随着文明社会的到来,这个过程被定格、凝固了起来,使我们看到今天的格局。C型主要分布于澳大利亚、伊利安、太平洋、西伯利亚;D型主要分布于青藏高原、日本、安达曼群岛;O型主要分布于东亚和东南亚;P型主要在蒙古高原和中亚草原,并低频存在于O型群体中。CD(注:即棕色人中基因类型)和OP(注:即黄色人种基因类型)的对立就揭示了远东地区的现代人发展历史中最主要的一个更替过程,这个过程就是我们这里要建立的假说,新石器的黄种人取代了旧石器的棕种人。
把单倍群的分布区块化地画在地图上,我们更容易看到,C型处于远东的南北两端,而中间地区,实际上也已非常低的频率存在着。D型则处于东西两端,同样在中部低频存在。对这种现象,可以有两种解释。其一,人群进入东亚以后,内部共有着C、D、O、P等类型,当进一步向周边扩张的时候,由于迁徙的人口较少,正好只有一小部分的CD个体来到了周边,发展成今天的样式。这就是一种瓶颈效应的解释。其二,CD的群体先来到远东地区,并且定居到了各个小区域,OP到达以后渐渐从中间向周边取代CD。这是群体更替的解释。对这两种解释的取舍,仅仅从群体类型的格局上是无法判断的,还必须借助于其它的证据。从Y染色体的STR标记,我们可以计算出各个单倍群的发生年代。把现在得到的所有有关个体都集中起来168Journalof National Dr. Sun Yat-sen MemorialHall计算,我们得到C、D的总年代为约95000年,而O、P的总年代却是约53000年。如果是瓶颈效应产生的格局,那么CD和OP的年龄应该是一样的,因为进入时他们是一个群体。而取代说才会是有不同的年代,CD的年龄要比OP大得多。所以,这基本上可以肯定是群体更替造成的,是OP正在替代CD。
我们把CD确定为棕色人种类型,把OP确定为黄色人种类型,是通过这两个人种的大量的群体分析得出的。在典型的棕色人种中,澳大利亚、伊利安和美拉尼西亚,最主要的类型是C,安达曼几乎全部是D。而黄色人种毋庸置疑,O和P始终是主流。所以OP替代CD就可以解释为使黄种人替代棕种人。 在母系的线粒体的分布中我们不仅能看到替代,也能看到交流。人类历史上,基本是母系的交流要多于父系的交流。就替代而言,同样的,线粒体单倍群M*和N*被ABCDFG等挤压在周边。在东亚大陆上,主要的是B、F,G、D、M7、M8。由于交流的大量存在,替代的格局不如Y染色体明显。
群体更替的观点确立以后,还有一点需要深入分析。因为更替也有两种可能,一种是外来的更替,另一种是内源的更替,两者都会得到替代者比被替代者年轻的格局。因此,必须把几个人种放在一起分析,看看黄种人是在棕种人内部还是在外部。白种人和黑种人必然在远东人类活动的大规模影响之外,如果黄种人的替代是内源性的,也就是从早期的棕色人种内的一群爆发开来的,那么他势必距离棕种人更近一些,而距离其它人种更远一些。反之,则会距离棕色人种远一些,至少不会近。所幸我们现在有足够的材料来做这样的分析。当我们用单倍遗传标记把各地区的人种进行聚类分析,得到了非常有趣的构型。非洲黑人自然很早就开始分化成几个细支,并不断地进行着基因交流。走出非洲的群体中,棕色人种最先分化,远远地离开了白色和黄色人种。白色和黄色的分化则相当晚,几乎是与棕色人种分化年代的一半。问题已经相当清楚了,在远东替代棕色人种的黄色人种是从外部进入的,入口就是东南亚。
这种替代的过程是怎么进行的呢?一种可能是黄种人进来以后,与棕种人通婚交流,渐渐地由于遗传机理而占到了优势。另一种可能是黄种人把遭遇到的棕种人消灭,占据他们到达的地方。或者,可能是一种中间的形式,消灭和交流兼有。从Y染色体看,主要地区的棕色类型CD都非常少,除了波利尼西亚(Polynesia)的黄色人种中间,看似棕色的男性可能大部分被消灭了。从线粒体看,人种之间的差异也是相当大的,但是交流的迹象相对多得多。所以女性可能被保留得稍多一点。要进一步看这种交流的程度,还是要看双倍遗传的常染色体基因。我们来看一下远东特色的HgC基因的几种类型在远东各地的分布。
其中,1型在东亚大陆,2 型在太平洋岛屿,3 型在北亚,这些都属于黄种人。*型是根部的未区分类型,澳大利亚棕色人种中全部都是这一类,南亚和东南亚也不少,但是在其它地区则基本上没有出现。为了分清这四个亚型之间的关系,我们进一步观察它们的网络关系图。
由这个网络图观察,我们要推翻前面的1、2、3型属于黄种人的论点。在Hgc起源位置上的原始类型都存在于南亚和澳大利亚的棕色人种群体中,而黄种人的几种亚型都起源于棕色人种,另外黄种人也没有原始亚型。这说明,Hgc不是黄种人原有的基因;黄种人在与棕色人种接触的早期,交流得到了少量Hgc;黄种人的进一步扩张过程中,人口膨胀使Hgc基因突变成新的亚型,但是并没有与棕色人种进一步交流,所以没有原始类型。现在可以得出结论,黄种人进入远东时曾与棕色人种有过有限的交流,而扩张期间则基本上把遭遇的棕色人种消灭了(所以说原始人是野蛮的原始时代生存竞争是十分残酷的)。现在我们可以勾画棕色人种和黄色人种在远东地区的迁徙历史了.A.棕色人种大约十万年前走出非洲,至少在五万年之前已经来到东南亚;黄色人种则是大约五万年前的时候开始向东迁徙;B.大约四万年前,棕色人种已散布于东北亚到澳洲的广大地区,而黄色人种则进入了东南亚;C.大约三万年前,黄色人种在东南亚、印度尼西亚次大陆和中国南方进一步扩张,而棕色人种可能有一部分已经来到美洲;D.大约两万年前,黄色人种几乎征服了东亚和东南亚的大部分地区,使该地区的棕色人种锐减;E.大约一万年前,黄种人进入美洲,并成为美洲的主宰;F.在数千年内,黄种人向中亚、北欧、印度洋和太平洋诸岛扩张,东亚最偏远地区的棕色人种也渐渐与黄种人融合。
远东地区两个人种扩张和替代的过程示意图
——A约50000年前;B约40000年前;C约30000年前;D约20000年前;E约10000年前;F约500年前。
这是一个动态的替代过程,黄种人扩张的同时棕种人也在继续前进。替代过程中的征服和杀戮恐怕是难免的。这与两个人种社会文化上的严重差异有关。黄种人之所以能替代棕色人种,也可能与其社会文化的相对先进和工具的相对先进有关。直到殖民时代到来,大部分的棕色人种始终都使用粗糙的旧石器,没有陶器;而黄种人则很早就有陶器和新石器。可能东亚地区的旧石器时代没有后续的晚期智人考古文化大多属于棕色人种,而新石器及其祖先型的旧石器文化则可能都属于黄色人种。社会和器物的更替通过人种的更替实现了。
【棕色人种的扩张史推测】棕色人种早期详细的迁徙历史已很难考证,他们是进入远东最有可能的线路是沿着阿拉伯海的沿岸。早期的棕色人种应该是以采集渔猎为生的,而且最可能是沿海打鱼为生。这是一种比较容易的谋生手段,海岸滩涂总是可以找到一些高蛋白的食物。一方面他们因为沿海觅食而渐渐向东迁徙,散布到了非常广阔的地域;另一方面,滩涂海产也是有限的,人口不可能过度增长。他们最大的分布特征是大范围低密度,这严重限制了他们的生产力发展。因为他们没有生存压力,就没有生产革新的动力。
我们可以想象,最早的棕色人种拿着最简陋的旧石器和尖木叉,在印度洋岸边走着,寻觅着鱼虾蚌蛤。经过了一代又一代人的扩展,几万年后他们来到了南亚和东南亚。在海岸边,到处是一个个棕色人种的小村落。当时的海平面比现在要低好多,巺他(Sunda)群岛与印度支那半岛连成一片,成为一个很大的印度尼西亚半岛。澳大利亚和伊利安以及美拉尼西亚的许多岛屿也连在一起。棕色人种很快就散布到了这片地区。到海平面抬升以后,印度尼西亚半岛消失,澳大利亚和伊利安远离亚洲大陆,棕色东南亚的尼格利陀(Negrito)小黑人人种在澳洲这两块世外桃源般的大陆上居住到了现在。而印度尼西亚和菲律宾(Phillipines)的偏远山区也还有零零落落的棕色人种部落,他们被称为尼格利陀人(Negrito)。 在南亚地区,印度半岛上的居民达罗毗荼人(Dravidian)长期以来都是棕色人种,直到白种的雅利安人(Arian)入侵才使他们发生了混血。他们是所知的棕色人种中文化发展的最为先进的一支。缅甸以西的海中,棕色人种也留存在安达曼群岛上,并保持着非常纯的血统。实际上来到远东以后,一些部落必然发展出以狩猎为生的生活方式,因而进入大陆腹地。他们来到荒无人迹的地方,没有任何的竞争者。只要用简陋的工具就能获得维持生存的食物。棕色人种的食物必然不会非常充足,没有精湛的狩猎技术无法保证每一次都有收获,所以他们不浪费任何食物来源。从安达曼到伊利安,棕色人种始终保持着食用人类尸体的传统,并以此建立他们的道德标准。尽管到处流传的食人族的传说总是让人不寒而栗,但对于棕色人种来说这是自古流传下来的部落生存必需的准则。这也成为后来他们为黄种人所不容的原因之一。
棕色人种北上东亚大陆的历史可能比南下澳州要晚一些。来到澳洲可能是六万年前,而在东亚大陆散布开来恐怕是四万多年前的事。中国的旧石器时代晚期文化或许大多是棕色人种的,虽然他们肤色可能并不深,与南方的棕色人种在体质上也可能有较大差异。至少,他们应该看上去不太像黄种人,面部的凹凸感较强,头颅形态也不同于黄种人。我们这里讨论的始终是晚期智人,与早期智人无涉。到了今天,早期智人的来源成了古人类研究中的最大的疑点。直立人起源于非洲,晚期智人也起源于非洲,而早期智人是直立人进化而来的还是另有渊源却是个未解之谜。但至少欧洲的早期智人源于非洲。 华夏地区的旧石器时代晚期人类遗骸有柳江人、山顶洞人、资阳人、札莱诺尔人等,他们的形态差异颇大。颜誾的观点比较客观,他指出柳江人比其它人更接近黄色人种。山顶洞人和资阳人很相似,有观点认为他们相对最接近虾夷人(Ainu)。我比较倾向于这种观点,这说明在三万年前,黄种人已经到达了中国南方珠江流域,而棕色人种还在长江流域和黄河流域活动。当时的日本列岛还与朝鲜半岛连在一起,是人类最容易到达的地区。捕捞生活的棕色人种就这样沿着海岸向东北延伸,通过白令海湾顺势到了美洲。
棕色人种到达美洲目前还没有遗传证据,但是许多古人类遗骨和考古遗迹的发现指向这一论点。在亚马逊(Amazon)发现的早期人类遗骸与印第安人完全不同,而更接近于澳大利亚土著。下加利福尼亚的一个特殊部落的发现更揭示了一个体质、文化和传统完全不同于印第安人的人群的生活。据说这个类似澳大利亚人的部落被西班牙人带来的瘟疫灭绝了。棕色人种在三万年前到了美洲之后过了两万年,黄种人才开始到来。黄种人和棕种人对美洲的争夺战可能在美洲大陆广泛地开展着。中美洲古老的奥梅克(Olmec)文化留下了许多石刻头像,无论如何都没办法把他们与黄种人联系起来。如果棕色人种来过美洲,他们一定就是。
【黄色人种的扩张史推测】黄种人走向远东的路线应该与棕种人不同,他们更可能采用了一条陆路。从西亚,进入印度河流域,在南亚次大陆的北端经过,进入东南亚。初来乍到的黄种人还没有能力与棕色人种抗争。至少在南亚的时候,印度半岛南部生活着大量的棕种人,一小批的黄种人只能沿喜马拉雅山南麓匆匆通过。这期间也融入了少量棕色人种的基因。在东南亚的密林里,黄种人发展壮大了起来,渐渐占据了优势。也是在这一时期,黄种人分化成了沿海支和内陆支。
沿海支的黄种人最初在越南、广东一带形成,后来部分人南下马来和印度尼西亚,部分人则沿着东海大陆架北上。这些人(水注:沿海支)就是南岛语系(Austronesian)和侗台语系(Daic)的人群的祖先,就是民族学上所说的百越民族。这一支的人群携带着明显的Y染色体单倍群O1。内陆支的黄种人则形成于缅甸、云南一带,并长期居住于此。由于历史比沿海支稍长,渐渐形成了更多的基因突变。这群人(水注:内陆支)是南亚语系(Austro-Asiatic)的祖先,就是民族学上说的百濮民族。他们带的Y染色体特色单倍群为O2 和O3。 我们这里提到某个人群带有某种特色单倍群时,并不是说他们只有这种单倍群,而是说他们以此为主,还兼有其它类型。在这些群体中,的确是特色单倍群占到绝大多数。
当时的末次冰期不仅仅造成了海平面的下降,大陆边缘的延伸,另一个效应是当时的一些不高的山都是积雪冰封。所以长江流域和珠江流域之间的南岭居然成了一个不小的阻隔。这限制了黄种人向北的迁徙。当然,可能还是有一些人进入了北方,这些人可能携带了特色Y单倍群P。他们(注:在冰期末最先翻过冰雪进入中国北方的黄种人)与后来到来的黄种人混合以后形成了黄种人的北亚支,就是阿尔泰语系(Altaic)的祖先。
末次冰期结束使黄种人大量北上,这个时间在大约两万年到一万年前。我们可以看到北方的人种发生了明显的变化,半坡和宝鸡的新石器人种与甘肃史前古人种完全不同。黄种人的史前大北伐使中国大陆的人类历史发生了革命性的变化。不久,新石器产生了,并迅速传遍了各地的黄种人社群,农业文明的星星之火在各地的黄种人中燃烧了起来。这开创了一个全新的时代,社会发展的火车头被拉动了。
黄种人的北伐也是分两条路线进行的。东部的沿海支进入浙江、江苏、山东,形成了河姆渡—崧泽—良渚的先越文化和大汶口—龙山的先夷文化。这一支人群与南下的沿海支黄种人遗传上比较接近,所以古人类学家观察到大汶口文化的人种体质上与玻利尼西亚的南岛民族极为相似。夷越类群(注:沿海支北上,商人与此类群关系密切)在沿海一带的发展相当快,东北渤海周边的红山文化也可能与这一类群有关。农业、新石器、玉器和早期的中国哲学思想都可能先产生于这一族群中。
西部内陆支的先濮民族也几乎同时北上,他们是通过云南、贵州、四川这条路线北上的,并且至少分出了两支。一支沿着长江往下走,到了三峡地区和洞庭湖平原,发展出了城背溪—大溪—青龙泉的先蛮文化,形成以后的苗瑶民族(Hmong-Mien)。另一支(注:内陆支北上,和夏、周有关)沿着四川的几条大河北上,来到了黄河上游地区,发展出了半坡—仰韶的先羌文化,形成了以后的羌华民族(Sino-Tibetan)。我们计算北方的O型的年代时,基本上都得到一万年左右的数值。 北部的阿尔泰群与当时北上的两支黄种人广泛地发生了混血。至今可以明显地看到,东部混有较多的沿海支成分O1,中部混有较多的内陆支成分O3。在混血发生之前从北部支已经有一部分的人开始通过白令路桥向美洲跋涉,这部分人在行进过程中Y染色体从P变成了Q。一万多年前到了美洲以后,他们(大陆北部黄种人)迅速地向南扩张,在千年时间内遍及了南北美洲。
从Y染色体看黄种人的迁徙,Underhill画出了一系列的线路图。他并没有在路线的细节上画得很详细,因为有些细节还不够清楚。比如P北上的路线,究竟是波斯还是中国,当时还不很清楚。现在在中国南方发现很低频的P 的原始类型,说明P更可能是通过中国大陆北上的。
黄种人来到北方以后,对当地的棕种人进行了杀戮和驱逐。但是这个过程并不是很快就完成的。棕种人已经在北方生活了这么长时间,并继续在许多偏远的山区躲藏了很长时间。直到商代的人骨材料中,我们还可以发现可能是棕色人种的成分,而且为数不少。只是他们依然是黄种人杀戮的对象。在殷墟的祭祀坑里有大量的异族俘虏的头骨,其中很大一部分可能是棕色人种。 这个替换的过程在中国进行到了非常晚的时期。唐代昆仑奴的传说不知道是否与此有关。据地方史料记载,清代广东官宦家庭中还有过豢养“小黑人”的案例。在台湾,最早到达的南岛语系原住民有着灭绝“矮黑人”的传说。这些零零星星的记忆都指向一点,黄种人对棕种人进行了种族灭绝。所幸的是,在菲律宾和印度尼西亚,这个棕色人种的分支——“矮黑人”还是存在的。我们无需对此忏悔,这是那个社会阶段必然要发生的事件。 在原始社会,与本种群有竞争关系的其它种群就是敌人,只有你死我活的结局。要么黄种人消灭棕种人,要么棕种人吃光黄种人。在这个长期的拉锯战中没有其它的结局。而当黄种人文化上先进于棕种人的时候,结局就只有一个了。那就是黄种人成了远东唯一的主宰。 当历史进入了阶级社会,情况就发生了变化,奴隶的需求使得黄种人对遭遇到的棕种人不再灭杀。那段时期黄种人才到达的边缘地区,比如青藏高原、日本列岛,棕色人种的成分被大量地保留了下来。而南岛语系的族群向两个大洋拓展的过程中,甚至避开了棕色人种居住的区域。
【两个人种的交融地区】日本列岛在棕色人种到来时还可以通过大陆架很容易地到达,而当黄色人种来到东北亚时,已成为茫茫大洋相隔的海岛,是黄种人传说中的蓬莱仙岛。所以长期以来,岛上居住的绳文人一直是与大陆不同的人种。关于绳文人的来源,日本研究者的大胆猜想很多。有一点是公认的,现代北海道的虾夷人和种子岛的部分居民应该是绳文人的后代。那么根据虾夷人的遗传结构,我们能够了解绳文人的来源。很幸运,虾夷人的Y染色体给出了明确的结论,他们大多是D,还有部分的C。他们必然是早期的棕色人种的后代。 日本的黄种人——弥生人,是中国的春秋战国时期迁入的,进入口是九州岛岛。他们向本州岛及更东北方向扩张的过程中,对旧石器的绳文人进行了征服和奴役。势必造成了绳文人比例的下降。但是到1800年,我们可以看到本州岛岛的北部开始到库页岛,都还是虾夷人的领地。 从本各地的遗传结构看,棕色人种的遗传比例虽然是从西往东递减,但各地的该种成分都不在少数。所以日本人总体上应该算是两个人种的混血区域。 虾夷人(Ainu)及其1800年分布区 对于虾夷人奇特体质,以前的人种学家猜想很多,最后有人认为这是一个处于黄种人和白种人之间的过渡种群。这实在是一种匪夷所思的观点,他们居然与远在地球另一面的白种人有关。其实他们的体质形态独特性说明,棕色人种经过漫长的历史发展,已然形成了几种不同的体质,至少澳洲土著、尼格利陀人和虾夷人是有区别的。 青藏高原的情况要复杂得多。藏族是一个很晚才形成的民族。他们是秦汉时期开始向高原迁徙的羌族中的两支——发羌(黄色人种)和唐旄(棕色人种)。在进入高原后,与当地的雅砻土著发生了混血。在藏族起源的神话传说中,雅砻土著被转化成了丑陋的猕猴,羌人则是仙女,两者婚配繁衍出了藏族。其实雅砻土著属于棕色人种,在羌人的审美观念中,他们的确是非常丑陋的。他们的混血使羌人很快适应了高原的环境。也使青藏高原的历史进入了一个全新的时期。我们在研究现在的藏族人的遗传结构时,还是能看到大量的D型Y染色体。而观察藏族的面貌特征,甚至在同一个村子中也会看到两种截然不同的样式。有的人是挺拔的羌人的面容,有的人则是敦圆的棕色人种面貌,肤色的差异也很大。
当我们在青藏高原进一步扩大调查范围,研究氐羌的其它民族,更有趣的现象出现了。曾经在中国南北朝的历史舞台上叱咤风云的氐族,现在仅存于四川和甘肃边界地区,有着惊人的Y染色体遗传结构——全部的D 型。难道他们也是棕色土著的后代? 我们恍然大悟,原来在汉藏语中,“氐”这个词,从来就是“底层”的意思。羌人来到青藏高原,把原本就在这里的土著居民称为“氐族”,因为他们“原抵”就在那里。我们再看羌族的结构,他们也含有部分的D型,但还是以O2为主。再看华族,就几乎不含有D型了。所以,“氐—羌—华”原初大概就是这么分化的。 而历史上对西戎民族的认识,一开始就有氐和羌的不同概念。我们进一步计算当地的D和O的年代,发现D至少在四万年以上,而O却最多纔有一万年历史。青藏高原上的D型成分与安达曼群岛棕色人种的D型应该是有关系的。 氐羌民族的几种面貌——A 白马氐族妇女(摘自注23);B 羌族妇女;C 黄种特征康巴男子;D 棕种特征的康巴妇女;E牦牛羌纳西族妇女 在东南亚的中南半岛,这样的遗传成分也是存在的,并且在各个民族中的成分比例不同。东南亚是世界上民族结构最复杂的地区之一,南亚语系的百濮民族是最古老的黄种居民。由于生活于热带,他们的肤色很深,比如佤族就是我国肤色最深的一个民族。但是有些民族的肤色就深得过份了,比如泰国和马来西亚边境上的土著Senoi族,再加上他们其它一些特别体征,不得不让人怀疑他们是否含有大量棕色成分。可惜他们的遗传结构至今还没有调查过。【转自何新博客:http://blog.hexun.com/songkoro/1033278/viewarti】
叶智彰:人类起源与中华民族的形成
一、灵长类动物与人类
灵长类在分类学上称为灵长目。这个目是生物中进化程度最高的一个目,它包括原猴类、猴类、猿类和人类,如果用分类学方法细分的话,可以分得非常详细,计有18个科161属,其中已绝灭7个科99属,现生者有11科62属222个种。
由于高度进化,人类摆脱了自然界的控制,反过来用自己的智慧去利用和改造自然界,故称为智慧生物。因此在灵长类学研究中把灵长类区分为人类和灵长类动物,灵长类动物统称为非人灵长类。灵长类学研究内容非常广泛,现存种类统称为现生灵长类,古代已绝灭的种类称为古灵长类,又称化石灵长类。研究人类本身的科学称为人类学,研究猿人进化成现代人类的科学称为古人类学。
与人类遗传距离最接近的灵长类动物就是猿类,猿类又称类人猿。它包括属于小型猿类的长臂猿和属于大型猿类的猩猩、大猩猩和黑猩猩。黑猩猩与人的遗传距离最近,2003年美国学者古德曼等人研究分析了负责构造蛋白质的那部分基因,在这部分关键基因组中,人类与黑猩猩有99.4%完全一样。换句话说;使我们成为人的东西可能就藏在O.6%的基因组中。
还有一个有趣的话题就是野人问题。近些年来,多处传闻存在野人,尤其湖北省的神农架地区传说更多,被描述成介于现代大型猿类与现代人类之间的高等灵长类动物。有人从良好的愿望出发,希望能从物种进化角度填补人类进化的空白,但这个问题很少有本专业的学者热心关注。有几位来访的日本学者问我,到底神农架存在野人的可能性大还是不存在的可能性大。我的答复是不存在的可能性大,理由很简单,作为高等灵长类动物,可以说毫无例外是社会性动物,过着群体生活。据称,传闻中的野人一般是单个个体,如此少量的个体如确实存在,其物种是难免绝灭的。反过来说,今天几乎到处都有人类的足迹,若存在野人群体,至今未被发现也是难以想象的。
二、人类起源问题
约在800万年前,人类和黑猩猩从他们的共同祖先分离,一支发展为现今的黑猩猩,另一支经猿人阶段发展成现代人类。在这段历史长河中,黑猩猩的变化较小,而人类的进化则经历了大的飞跃,从四足型灵长类进化成直立行走手足分工的两足型灵长类,其大脑得到飞跃式的发展,成为地球上唯一的智慧生物。咸了控制和改变大自然的主人和地球上的万物之主,发展成统治整个星球的物种。
人类起源研究存在着“单一起源论”和“连续进化论”两种论点。前者认为人类只起源于一个地区;后者认为非洲、欧洲、亚洲的猿人各自进化成现在的黑种人、白种人和黄种人。科学家1974年在埃塞俄比亚发现一具保存完好的女性古猿骨骼,取名“露西”。她被单一起源论者认为是人类共同祖先,约生活在300多万年前的东非。2002年,法国科学家米歇尔.布吕内在撒哈拉乍得的沙丘中发现一个距今600万~700万年的人种:撒哈拉乍得种。所以,在谁是最古老的原始人这个问题上还存在激烈的争论。在判定古人类化石的年代上,古人类学家和遗传学家存在很大的差异。后者根据只存在母性遗传的线粒体DNA硷基替换速率推算出“露西”生活在距今30多万年而不是300多万年。
国际上现今较多学者尤其是遗传学者支持单一起源论,认为约在13万年前古人类走出非洲,沿海岸线经南亚和东南亚岛链到达澳洲,然后沿原路线返回到现今的伊朗,分为两支,一支向北迁移,形成高加索人种(白种),另一支沿海岸线东进,在我国南方广东一带登陆(约在6~1.8万年前);另一说是约在同一时期同时经广东和云南进入我国南方,在中华大地形成蒙古人种(黄种)。而留在非洲和到澳洲的人群形成了尼格罗一澳大利亚人种或称尼格罗人种(黑种)。当时的海平面比现在低几十米,岛链之间有陆桥相连。据印度生物学家2005年的研究,印度安达曼一尼科巴群岛上的两个原始部落可能是至少5万年前第一批从非洲向外迁移的人类的直接后裔。DNA研究表明,这两个部落是“世界上最早人类的后裔,也是第一批走出非洲的现代人类”。这些部落的体貌特征与非洲人相似。由母系遗传,存在于人类每个细胞之中的线粒体DNA可以一直上溯到生活在约15万至20万年前的同一位母系祖先身上。细胞和分子生物中心主任拉勒吉.格教授说,被研究的原始部落居民的DNA与那位母系祖先所谓非洲源头的基因极其接近。因此他们很可能是5万至7万年前通过海路从非洲迁移到此,并从此就一直在安达曼群岛过着“基因隔离”的生活。
在人类起源问题上,较多的学者尤其是遗传学家支持单一起源论。人类起源于非洲的依据,一是在非洲发现的古人类化石其生活年代最古远;二是根据遗传多样性。不同人类群体的基因遗传多样性各不相同,科学家们发现,黑人比其他人类群体——白种人、黄种人等拥有更丰富的遗传多样性。也就是说,黑人的基因种类更多。比如由基因导致的血型差异,黑人中A、B、AB、O血型都普遍存在,但欧洲人中B血型非常少见。基因研究发现,黄种人、白种人的基因表现形式在黑人中都存在,反之,一些存在于黑人中的基因却在其他人种身上没有发现。据此,得出了黑人是现代人祖先的结论。比如,一个地区的人类有1O种基因,但并不是存在于每一个人的身上。因此,当一部分人迁移出去时,并不能带走所有的基因种类,只能带走几种。当这些人繁衍生息,其后代只继承了这一部分基因,遗传多样性自然就比祖先地区人类少了。
对人类线粒体DNA的研究显示,今天活在世界上的所有人都可以把自己的祖先追溯到大约15万年以前生活在非洲的智人。当时,世界上还有两种原始人:生活在欧洲的尼安德特人和生活在亚洲的直立人。但在智人从非洲迁移到其他地方以后不久,这两个人种都消失了。多年来,科学家一直没法解释尼安德特人和直立人的消失。他们提出从战争到智人从非洲带来的奇异病毒等各种理由。但这两个人种灭绝的原因可能要复杂得多。
三、中华民族的形成
我国的人类学研究,人类学家一般持连续进化论观点,而遗传学家则持单一起源论观点。但是,不论从体质人类学的研究结果,还是从分子遗传学研究结果来看,蒙古人种(黄种)均可分为南北两大类,即蒙古人种南方类群和蒙古人种北方类群。也就是说两种不同论点的学者所得出的研究结果是相同的。在体质人类学研究方面,张振标等(1988)研究了现代中国人(包括汉族和少数民族)的体质特征。结果表明,现代中国人在体质人类学上表现出两种类型:以长江为界,长江以北的居民属北部类型,而长江以南的居民则表现为南部类型。用我国新石器时代颅骨聚类分析的结果也同样表明,南北两大类特征明显。他据此指出,从人种起源角度看,现代中国人的这两个体质类型形成的并存不是源于不同人种的混杂,而是来源于同一人种——黄种人。并指出,导致两个体质类型形成的根源可以追溯至中国晚期智人。北部类型可能是以山顶洞人为代表的北部地区晚期智人发展而来,南部类型可能是以柳江人为代表的南部地区晚期智人发展而来的。林圣龙(1987)根据早期人类化石的形态序列和时间序列认为,中更新世早期的人类,从长江流域向北越过秦岭山脉,逐渐扩散到黄河流域。他推测蒙古人种两大亚类的形成可能有几十万年的历史。
分子遗传学研究的结果也异曲同工地获得了上述结论。赵桐茂等对现代中国人免疫球蛋白同种异型的分析(1987,1991),和运用人类白细胞抗原等频率计算人群间的遗传距离(1984),以及杜若甫等对中国人群间遗传距离的研究(1983,1998)等结果都表明,大致以长江为界,中国人明显地分为南北两大类群。北方少数民族向北方汉族聚类,而南方少数民族向南方汉族聚类。也就是说,南北汉族之间和南北少数民族之间遗传距离大,而北方汉族与北方少数民族之间和南方汉族与南方少数民族之间的遗传距离小。藏族向北方类群聚类,是因为北方人群经青海向西藏扩散的结果。
南北类群的形成是由于长江地理隔离所致。在古代南北人群因地理隔离缺少基因交换而形成了遗传学上的差异。根据免疫球蛋白同种异型Gm因子在世界范围的分布,人们认为北方类型的蒙古人种后来扩散到朝鲜和日本,一部分通过白令海峡进入美洲。所以北方类群包括中国北方人和东北亚各地居民如日本人、朝鲜人、蒙古人及俄罗斯亚洲部分的少数民族,此外还有美洲的印地安人等。而南方类型的蒙古人群向东南亚各地迁移,因此,南方类群包括中国南方人和东南亚各地居民如泰国人、印度尼西亚人和菲律宾人等。以前所谓的马来人种(棕种),应属蒙古人种(黄种)南方类群。根据最近美国学者乔迪.海伊对美洲蒙古人种DNA的研究,(2005),美洲的蒙古人群是在约1.2~1.4万年前的冰河期穿过白令海峡到美洲的,当时约有70人到美洲,现在美洲的蒙古人群如印地安等都是这70人的后裔。而以前认为这批蒙古人群是在约2.5万年前到美洲的。中国处于两大蒙古人群的原居栖地,因此Gm单体型频率分布呈现“混合状态”。居住在我国西北地区的维吾尔族、哈萨克族、东乡族和回族具有Gm3;5单体型,这提示这些民族混有高加索人种血缘,其中,维吾尔族、哈萨克族和东乡族的混合程度都大于25%。
体质人类学研究和分子遗传学研究结果均表明汉族不是一个同源的群体。居住在北方的少数民族和北方汉族在同一个集群,居住在南方的少数民族和南方汉族在同一个群体。这说明从种族关系上看,南方和北方汉族之间的差异,远远大于汉族和少数民族之间的差异。作为一个国家,北方和南方两大类群虽然有较多的血缘交融和基因流动,但各自的体质人类学特征和分子遗传学特征并未消失。
在中国历史上,中原汉人南迁,到中国的南方繁衍生息,形成了几个大的民系,几大方言区,如客家民系、广府民系、河洛民系等。同是中原汉人,同是讲北方方言,但到了南方不同地域以后,却形成了现今不同的方言。究其原因,很大程度上取决于当地原住民所使用的语言。现今南方汉族的几大民系都称自己的祖先是中原汉人,现在要讨论的是在血缘上是北方中原汉人为主呢,还是南方原住民为主。这两者之间虽然有较多的血缘交融或基因流动,但各自的人类学和遗传学特征并未消失。大量这方面的研究表明,现在的中国南方人属于蒙古人种南方类群,这只能认为迁到南方的北方中原汉人占少数,只是将当时较先进的中原汉文化带到南方,同化了当时占多数的南方原住民。
有趣的是,根据复旦大学文波等人的研究(2004)显示:汉文化的扩散的确伴随着大规模人口迁徙,但这场由北向南的人口迁徙中,占主导地位的是男性。研究人员测定和分析了来自南北28个地区有一定代表性的汉族人群的Y染色体非重组区和线粒体DNA,结果发现,南方汉族与北方汉族的Y染色体类型及频率分布非常相近。母系方面,北方汉族与南方汉族的线粒体类型分布非常不同。这可以说形成了南北汉族“同父异母”现象。
以上的研究结果可以理解为,古代较大规模的中原汉人南迁,多数为男性,迁到南方与南方女性通婚并繁衍生息,形成了南北汉族“同父异母”现象。有史料记载的这一个例子就是秦始皇派尉屠睢率50万大军驻岭南。这支庞大的部队大都是男性,他们在秦亡后没有北返,继续留在当地,这是大批汉人南下之始。汉武帝时,发大兵南下平定南越,然后在秦代南疆三郡的基础上设置九郡。同样,这批大兵也没有北返。这样的向南扩张属于政治和军事上的征服过程,其直接结果是汉文化的渗透和同化。后来,从汉末到东晋,受黄巾起义和永嘉之乱影响,大批中原汉人南迁。在中原汉人南迁的潮流中,最具代表性的当属南北朝时,北方汉人为躲避战乱而大量南迁。这次南迁对中国人口版图的划分具有重大意义。当时南迁人口的数目,据说占南方人口的六分之一,当时东晋的人口为540万,以此计算,当时南迁人数至少有90万,这些移民看来同样男性占多数。最后一点推论就是,当时较流行的一夫多妻制进一步促进了南北汉族在遗传特征上“同父异母”现象的形成。
综上所述,人类起源于非洲,现今世界上存在的黑种人、白种人和黄种人在动物分类学上属于同一物种的3个不同亚种。中国人的祖先先在中国南方繁衍生息,后来扩展到北方。由于长江形成的地理隔离,南北人群在遗传上缺乏基因交流,形成了具有不同遗传特征的蒙古人种(黄种)南方类群和北方类群。历史上虽然发生过较大规模的北方中原汉人南迁,但在遗传上并没有取代南方地区以前存在的人群。中原汉人南迁带动了汉文化的扩散,将当时较先进的中原汉文化带到南方,并将南方人群同化,使其成为汉族的一部分,但文化上的同化无法改变南方人身上的遗传基因。所以,汉族只是文化上而非血缘上的完整群体,南北汉族在遗传学上属于具有不同遗传特征的不同种族,由于文化相同而形成了同一的民族。(叶智彰,现任中国科学院昆明动物研究所任职;摘自2006年第1期《云南科技管理》)
附录:陕西神木突现宏大古城史书无记载
(新快报记者刘虎报道)陕西神木发现我国史前最大城址,史书中尚未找到针对性的文字记载。因发掘工作浩大,可能耗时十数年或数十年。这座硕大无比的史前古城正在改写中国上古史及中华文明的起源。
神木县位于黄河中游、长城沿线;境内丘陵密布,沟壑纵横;草滩绵延,沙梁翻滚;是一块荒凉贫瘠的土地,历史上多次发生过战争,狼烟四起,只有饥民逃荒,不见富户荣归故里。因此,尽管它是陕西省最大的县,总面积7706平方公里,但却人烟稀少。直到二十多年前勘探人员在这里找到了世界级的“神府-东胜”煤田,滚滚煤海才让神木人过上了富裕生活。
要说这里有一座我国史前时期最大的古城,似乎像是一个笑话。不过,若是你来到黄河支流秃尾河及其支流洞川沟交汇处的山梁顶部实地,看到已经清理发掘出的一个面朝东方、居高临下、位置险要、占地2500平方米的城门遗址,大概就要扭转看法了。
2011年由省、市、县三家文博机构组成联合考古队,对石峁遗址进行了区域系统调查,发现了一处规模宏大的石砌城址。2012年,经国家文物局批准,正式对石峁遗址进行了重点发掘及复查,并确认了石峁城址由“皇城台”、内城、外城组成。
内城墙体残长2千米,面积约235万平方米;外城墙体残长2.84千米,面积约420万平方米。其规模远大于年代相近的浙江良渚遗址(300多万平方米)和山西陶寺遗址(270万平方米),是目前所知我国规模最大的新石器晚期城址。
华夏第一门
发掘工作从2012年5月至11月。主持发掘的陕西省考古研究院院长助理孙周勇说,为了弄清楚石峁城址到底建于何年代,考古队当年重点发掘东门。这是我国迄今考古发掘的最早石城门,其结构异常复杂。发掘显示:它主要由“外瓮城”、两座包石夯土墩台、曲尺形“内瓮城、门塾”等部分组成,这些设施以宽约9米的“”形门道连接,总面积约2500平方米。
东门遗址属于整座城的地势最高处。石墙墙体宽约2.5米,保存最好处高出龙山晚期地面4米。整个外城的东南角与其相连,地势开阔,位置险要,堪称整个古城的枢纽地带。曾在考古现场考察过的四川省考古研究院院长高大伦认为,从总体上讲,石峁古城打破了中国夯土筑城的传统认知,其东门遗址无疑是目前考古发掘所见最早的石城门,其功能和结构的复杂性尚有待探讨,其在黄河文明史、甚至人类文明史上的意义都是难以估量的,在中华文化史上的地位堪称“华夏第一门”。
48个人头骨
此次考古发掘,首次考古发现7件玉器,其中完整的6件,主要是玉铲、玉璜等,最长的玉铲有18厘米长。不过,这些玉器出土的地点都十分奇特——在城墙里。石峁考古队队员邵晶此前听当地老乡讲,上世纪九十年代开始,大批玉器流散出来,出土的地点是就石头墙里面。“我们当时还有些怀疑。后来发掘证实老乡说的是对的,我们这些玉器出土有两种情况,一种是在倾斜的石墙里面,第二种就是在倒塌的石墙堆积里面。”考古队认为,这也为流散在海内外的石峁玉器找到了确切的时代依据和文化背景,证实其有4000年历史。
考古中另一个出乎意料的发现,是遗址下层地面下出土了48个人头骨——考古队找到了集中埋置人头骨的遗迹两处。这两处人头骨摆放方式似有一定规律,但没有明显的挖坑放置迹象,每处各有24个头骨。经初步鉴定,这些头骨以年轻女性居多,部分头骨有明显的砍斫痕迹,个别还有灼烧迹象。考古人员推测:这些头骨可能用于4300年前城墙修建时的奠基或祭祀活动。
改写中华文明的起源
2012年10月,中国考古学会、国家文物局、陕西省文物局、中国社科院考古研究所、国家博物馆等40余位考古专家,对神木县石峁遗址发掘现场进行了联合考察。
“我看了以后感觉十分震撼!”中国考古学会理事长张忠培先生认为,该遗址的价值不是今天就能完全认识到位的。河套地区规模最大的石头城、用玉器来表现文化、黄河腹地二里头文化之外另一个重要遗址,这些都足以体现了遗址的重要性。
著名考古学家李伯谦先生认为,石峁遗址的发现,为中国文明起源形成的多元性和发展过程提供了全新的研究资料。陕西省考古研究院认为,这4000多年前的史前最大城址,门址、石城墙、墩台、“门塾”、内外“瓮城”等重要遗迹体量巨大、结构复杂、构筑技术先进;初步认定石峁城址最早当修建于龙山中期或略晚,兴盛于龙山晚期,夏时期毁弃,属于我国北方地区一个超大型中心聚落。
对话三位考古学家
“这古城,我们有太多不知道”
●对话人物:陕西省考古研究院院长助理、神木石峁遗址考古队领队孙周勇陕西省考古研究院研究员、商周研究部前部长张天恩
中国社会科学院考古所研究员、夏商周考古研究室主任兼二里头工作队队长许宏
玉器是有意埋进城墙里的
记者:石峁遗址的发现比较久了。为什么选择在2012年启动发掘?张天恩:2011年对遗址做了一个全面的勘查,发现这个城非常大,400多万平方米,龙山时期最大的城。过去文物普查的时候知道有城,但并没有想到规模有这么大,没有引起足够的重视。发现后,2012年就动工了。
记者:发掘出的玉器是建城时埋进墙里去的吗?还是后面放进去的?孙周勇:具体原因目前还不能解释,但可以讲我们观察到的现象。玉器是有意埋藏进去的,在修建过程中。这个很难理解,很可能是跟祭祀礼仪宗教活动有关。
记者:埋那么多人的头骨也跟祭祀有关?孙周勇:对,应该也跟造城时的祭祀有关。这座城,证实在公元前2000年-2500年的时候,北方有一个很大的集团中心。至于它是谁的,我们还没法说。过去,我们以为北方不是那么发达,是蛮荒之地。但现在,知道那时有类似现在有北京、有西安、有广州这样的格局。
它存在于华夏形成之前
记者:现在知道这座城有几个城门?孙周勇:内城和外城各有东城门,其他的不好确定。外城的东城门在全城最高处,有防御性的作用,便于瞭望观察。
记者:在石峁古城存续期间,那块土地属于谁的势力范围?是不是华夏之域?许宏:沒有文字记载,不知道是哪个邦国的。石峁城址应该是一处区域性的中心吧。孙周勇:它存在于华夏形成之前,也不在后来华夏的区域。张天恩:它属于哪个部族,没有说法,找不到记载——我翻的所有的书上都没有。这个考古应该是中国北方地区文明最重要的发现。
记者:如此宏大的规模,您认为它可能是一个什么样的城市?是不是一座都城?孙周勇:“都城”的概念,那时不一定有。我倾向于酋帮,就是部落,大的集团,这个城址是大的势力范围控制的一个中心。许宏:石峁是当时那一带最大的城址,如有都城,它就应该是。张天恩:地域性都城,相当于中原地区夏朝的都城。它就是龙山时期最大的城。
记者:古城废弃原因是什么?张天恩:也许是因为战争,或者是人类活动中心的转移,又或者是因为气候环境改变得很恶劣,越来越冷,不适宜人的居住。现在文献一点线索都没有,期待靠发掘来解决,找到更合理的说法。
重庆发现人类最早艺术品人类非洲起源说受到挑战(新华网)
重庆商报12月20日消息:去年5月,当地群众报告,天坑地缝景区洞穴中发现几处化石点。为了抢救和保护文物点,奉节县风景管理局和文化旅游局邀请中国科学院古脊椎动物与古人类研究所研究员黄万波前往现场。黄万波发现,这些洞穴是史前人类穴居过的山洞,古人可能在洞中居住了上万年。天坑地缝地区10个洞穴中,他发现了8处古生物化石点和大量哺乳动物化石。
发现有图案的化石
去年9月上旬,黄万波教授再次率领科考人员,在天坑地缝周边进行抢救性发掘。经过一个月的野外工作,在云雾乡海拔高度约1100米的1号和2号洞内,发现了巨羊、硕箭猪、双角犀、剑齿象、大熊猫等30余种哺乳动物化石。同时,在1号洞离洞口50米处的堆积层,还发现许多旧石器和1枚古人牙化石。其中特别引人注目的是,在人牙的旁边,有排列整齐的一对完整的、长达2米多的剑齿象门牙化石。在门牙的尖端部表面有若干明显的痕迹,其中几条深而弯曲,像伸展的树枝,横断面呈U型。
黄万波称,史前时期兽骨或兽牙上的痕迹,产生的因素有6个方面:动物(啃齿类或食肉动物)的咬啃;大型哺乳动物踩踏;植物根茎腐蚀;自然营力作用(包括物理风化、化学分解、地层滑动等);动物自己磨蹭;史前人类刻划或砍砸。经史前考古、古生物学者和动物专家观察研究,排除了产生这种痕迹是自然营力、动物啃咬、大象生前自己所蹭等因素,认为产生这种痕迹的因素,最大可能是中更新世“奉节人”所为。
确认为最早艺术品
去年11月19日,黄万波在渝就发掘情况召开新闻发布会,通报了这一令人振奋的好消息。他对我市和中央驻渝各媒体记者称,刻痕是人类艺术最早的萌芽。奉节发现的中更新世晚期刻痕与欧洲曾发现的一种史前人类刻痕非常相似。对此次发现,他们还将采用电子显微镜扫描、压模对比等进行研究。如果进一步研究结果能得以印证,将标志着青藏高原东部的长江三峡地区出现了人类最早的刻痕艺术。根据随同出土的各种动物化石群推算,人牙是距今数十万年前中更新世古“奉节人”的化石,将为“三峡是人类重要起源地之一”提供实物证据。具体年代尚待进一步测定。
经中科院和英国大英博物馆进行严格的科学鉴定研究,这些痕迹最终被认定为鸟头图案,并确信为史前人类刻划所致。其年代经中科院地质研究所和南京师范大学分别用考古铀系法进行科学测定,最近得出了非常接近的结论:一为15万年,一为13万多年,都在14万年上下。它比目前发现最早的史前人类刻划在一块南非矿石——赭石上的图案还要早6万年,是人类最早的艺术品。
中国学者用年差挑战“非洲起源”说不完备_广州大洋网
(一)2002年末我国传出惊人的消息,现代中国人的祖先更可能在中国,而不是非洲。这是我国科学家用世界先进的地层年代测试方法,首次对埋藏柳江人头骨化石的地层进行年代测定后得出的结果。广西自然博物馆副研究员王頠(wěi)向新华社记者透露,他的研究小组通过对柳江人洞的堆积物进行详细研究,采用铀系测年法,首次对柳江人头骨化石的出土地点进行了系统的地层年代测定,结果表明,柳江人可能生活在距今7万年至13万年之间或更早,而不是原先所认识的不超过3万年。这意味着,具有解剖学上现代人特征的柳江人——现代中国人的祖先,比人类学家原先所认识的要更早就生活在华南地区。
这一结果使现代人“非洲起源”学说受到挑战,这一学说认为,现代中国人是3.5至8.9万年前从非洲迁移来的。王頠说,他们所采用的测年方法,突破了以前同位素碳-14测定法最多只能测到3万年的局限,而把可检测的最远时间延长到数十万年,这种方法目前在第4纪研究中,已被公认为是一种最为成熟、可靠的新方法。1958年在广西柳州市附近的柳江县一个洞穴内出土的柳江人化石,包括一个基本完整的头骨和部分体骨、肢骨,是迄今为止东亚地区发现的少有的基本完整的现代人的化石之一。对它的年代测定有助于理解现代人类起源——这一国际学术界普遍争论的焦点问题。“化石出土的地点现存8米多厚的堆积层,可分上、中、下3个单元层,我们在不同层面抽取钙板样品,在两个世界著名的铀系测年实验室——分别在南京师范大学和澳大利亚昆士兰大学进行测试,并进行地层学分析,发现原埋藏柳江人头骨化石的中层的年代介于距今7万年至13万年之间,或更早。”王頠说。地球上最早出现古人类是在至少50万年前,之后先后演化成能人、直立人、早期智人等阶段,大约10万年前进化成晚期智人,现在在地球上生活的人类属于晚期智人,或称为解剖学上的现代人。
尽管古人类起源于非洲在国际学术界的看法比较趋向一致,但对于现代人的起源问题却有激烈的争论。去年5月,美国《科学》杂志刊登了由中外科学家组成的一个研究小组获得的最新研究成果,他们通过基因研究认为,东亚人的祖先不是四五十万年前的北京人,而是来源于非洲的现代人,他们于3.5至8.9万年前从非洲迁移到东亚地区。“用基因研究的结果推测人类进化过程,无论如何是间接的,而来自化石的证据却是直接的。柳江人化石年代的测定直接证明了在7万年至13万年前或更早,柳江人就已经在华南地区生活了,而不可能是在这之后才从非洲迁移来,”王頠说。
中国最早的古人类化石是170万年前的元谋人牙化石,在中国出土的各时期的古人类化石都有共同特征,如铲形门齿、长方形眼眶、脸较为扁平等,这表明至少170万年以来,中国人的进化过程是连续不断的。但由于东亚地区缺少5万年至10万年前的人类化石证据,使这种连续性出现了缺环。基因研究者因此认为,东亚地区的古人类在这个时期灭绝了,非洲现代人迁移亚洲并取代原住人群便“顺理成章”。“我们的新发现弥补了这一缺环。在不久的将来,包括年代学在内的对中国南方古人类遗址的系统综合研究,有望揭示现代人类在东亚出现的年代和演化模式,”王頠说。中科院古脊椎动物与古人类研究所研究员黄慰文说:“除人类化石证据外,在中国出土的旧石器工具,其打制技术主要继承了其祖先的技术,这也说明了现代中国人更可能起源于中国。”中科院古脊椎动物与古人类研究所研究员侯亚梅说:“过去由于东亚地区缺乏现代人的化石证据,使现代人‘非洲起源’说在国际学术界得到广泛认同。柳江人化石的年代测定,使国际学术界重新把目光转向东亚。”
2003年末我国又传出惊人的消息,中国科学家和考古专家对萨拉乌苏文化遗址的最新研究表明,现代人多地区起源说获重大证据:7万年前就有“鄂尔多斯人”(即过去称呼的“河套人”)。著名人类学家、中国科学院院士吴新智就是主张多地区起源说的专家之一。但一直以来,“多地区起源说”未占主流,更多的学者倾向于“非洲起源说”。这就有一个前提,如果能在亚洲找到早于6万年前具有现代人体质特征的古人类化石,“非洲起源说”就难以成立了。中国科学家上世纪80年代用碳14同位素方法对萨拉乌苏遗址测定,距今大约3.5万年,沿用至今。但由于这种方法的测定范围不超过4万年,所以,3.5万年只说明萨拉乌苏遗址不晚于3.5万年。中国科学院寒区旱区环境与工程研究所研究员董光荣采用另一种同位素方法———热释光方法所获结果为距今7万年至15万年。不久前,中国地震局地震研究所尹功明用更加先进的光释光方法测定,也证明萨拉乌苏遗址的年代不晚于距今7万年。上述结果符合萨拉乌苏动物群和地层学观察对于遗址的判断。他们的研究发现得到中国科学院院士刘东生的充分肯定。因此,“鄂尔多斯人”的年代不晚于距今7万年的事实,将成为现代人多地区起源说的重要证据。
(二)中国部分科学家的这种不晚于距今7万年的年差的反驳,即如果能在亚洲找到早于6万年前具有现代人体质特征的古人类化石,“非洲起源说”就难以成立的证明;但如果其前提是20万年前生活在非洲的人类祖先,离开非洲分散到世界各地代替当地的土著居民,走的是陆路,这就不完备了;因为与陆路相对还有海路。
打开地图看吧!早在遥远的古代,印度洋上的阿拉伯海和孟加拉湾虽是阻碍东西方来往的一片难以逾越的水域,上面驮负着极其沉重的亚非大陆,索马里、阿拉伯、波斯和印度西部沙漠的风沙,紧贴着它的背脊尖声呼啸,一列列土黄色的沙丘一直伸展到海边,但早有古人开始考虑从海上开辟一条更直接便利的道路。阿拉伯人早在3000多年以前就顺着红海航行到了东非,还曾在波斯湾上航行过。世世代代在海上生活的结果,使他们逐渐发现了印度洋的秘密。他们发现,每年的11月到第二年的3月,风总是从东北方的大陆上吹来,拂动着海水向西南流去。这时的海上总是晴空万里,积云和雨水都很少。4月至11月则恰恰相反,出现西南风,驱赶着云涛和海流不断驰向东北方,海上的雷雨也比较多。
因此横渡阿拉伯海的办法说来也简单,只消掌握住风向和海流的变化规律,就能航船到远方的印度去。即夏、秋两季,可以利用西南风直航印度;冬、春季两行,再顺着东北风返回阿拉伯半岛和非洲。后来的人们也是依靠季风的帮助,十分顺利地建立起了和非洲、印度的联系,在海上铺平了一条连接东西方之路。我国从远古《山海经》诞生时开始,便流传了许多关于远方异国的传说。例如东方海外的黑齿国,那里的人们牙齿是黑色的,喜欢吃蛇,也能玩蛇;南方海外的灌头国的人嘴部突出,以捕鱼为生;西方海外的奇肱国人会捕捉各种飞鸟;北方海外的聂耳国的人,耳朵较长,住在海岛上,是猎虎的能手。这些有趣的传说,看起来仿佛都是充满了幻想色彩的荒诞神话,但是仔细加以分析,便会觉得其中的一些国度与印度洋及阿拉伯半岛和非洲上的许多地方相似。传说往往以现实为基础,其中有一些很可能是我国古人在海上的见闻实录,或在航途中从其他民族古人那里听来的,也许有一部分是真实的情况;。
总之,如果是20万年前生活在非洲的人类祖先,离开非洲有一部分迁移来到阿拉伯海的也门和阿曼海岸边生活,数万年间能发明一些用蒙着海豹和海象皮的小舟,或用芦苇捆扎成的小船,或者就是用独木舟,在缓缓漂浮木块的洋流水道上跟随着行进,又顺着风一桨又一桨地用力划行着,向着迷茫不清的海面驶去,虽到处都潜伏着不可捉摸的危险,但他们世世代代都生活在阿拉伯海岸,尽管许多古人曾经葬身在变幻无常和神秘莫测的海里,可是他们之中有人却汲取了更多的经验教训,勇敢地划着小皮舟或芦苇舟或独木舟,在东方的海面上越驶越远。而且,也许经验告诉他们,有一股海水从印度那边滔滔不绝地涌入,在这股海流下面,隐藏着数不清的鱼。这对捕鱼为生的海上古人来说,也是一种强烈的诱惑!终于有一次,他们驶行得比以往任何一次都远,逐渐驶入了那股夹藏着大量游鱼的暖流悄悄向东偏移,丝毫也不觉驶近了迎面而来的另一条陌生的印度海岸。后来又把他们送到了恒河流域,从印度到缅甸,缅甸到我国云南。如果遇上这一股是从印度通向阿拉伯半岛的洋流,反之亦然。
这项研究也是有根据的。曾有探险家研究了太平洋上的土阿莫图群岛的民族起源,又仔细察看了埃及一座大金字塔内壁画上的芦苇船图形;而与此相似的芦苇船,至今还在南美高原上的的的喀喀湖上应用,认为大洋不足以成为古人类往来的障碍,古人很可能就是乘坐芦苇小船漂航能到印度去。他们在青尼罗河发源地的达拉湖边砍了12吨芦苇,按照从金字塔里抄绘来的图纸捆扎了一条小船,大约一个月就漂航了3300公里。而关于独木舟,大约在17世纪末,英格兰中部兰开郡的马丁湖被排干了,当人们在开挖湖底的时候,无意中掘出了一只独木舟,接着又是一只……,就这样一共挖出了8只。最初人们以为这几只独木舟不过是英国古代原始人类的遗物,但是经一位名叫李依的学者仔细研究和比较分析这些独木舟的式样和大小,发现竟和当时美洲印第安人使用的没有什么不同。如果鉴定没有错,这唯一的可能性只能是古代的美洲印第安人曾经到达过英国。难道古代的印第安人就是凭借这种原始的独木舟漂过辽阔的大西洋到英国来?其实要回答这个问题并不难,从美洲中部炎热的墨西哥湾,也有一股宽达几个公里的墨西哥湾流,以每昼夜150公里的流速偏向北西,流到英伦三岛的西岸,然后再向北流到挪威的海面,最后消失在北极圈里。这股巨大的海上“河流”曾卷带着一些美洲的热带树木流到北欧沿岸,给古人以启发。独木舟漂洋过海虽然充满了危险,但是在顺利的情况下,却不是不可以成功的。
千百年来,不知有多少迷航的独木舟在漂洋越海的途中被大海所吞噬,真正到达印度海岸的只是其中很少的一部分。有趣的是,在与大海完全隔绝的四川各地区考古,都发现有一种像船的棺材悬在悬崖上,而并非发现于中国东南的海边。很可能,5000多年前四川还是内陆盆塞海,那些侥幸逃生的人类的非洲祖先,经阿拉伯海的洋流乘独木舟之类的工具到印度,从印度到缅甸,缅甸到云南,云南到广西,北至内蒙古,特别是进入中国后,就迁居到水草丰美、适宜于渔猎生活的四川内陆盆塞海,并按照曾在阿拉伯海湾生活的方式制造了这些像船的悬棺的独木舟。在四川省盐亭县天垣乡盘垭村发掘出的“盘古王表”和盘古王退位后南迁的传说,以及在四川发掘出的三星堆、金沙等古遗址发现的远古文明,和巴比伦古苏美尔人是蜀人等,也为这种东西方往来的远古交流提供了证据。因此,20万年前生活在非洲的人类祖先迁徙到中国,走的是海路,就只需10到15万年,用年差挑战“非洲起源”说难于成立。因为柳江人和鄂尔多斯人的时间测定在10到15万年区域,说明可能是从海路来中国的。
(三)通过我国著名冰川学家韩同林等人的考察研究,第四纪大冰川期,中国已成地球大冰球的一部分。如果设想在约100多万年前,全球各大洲虽都有直立人,但为避惧严寒,他们中的一些优势群体都大迁徙到了非洲,约20万年前又从非洲走出,这样既能回答各地的人类起源化石发现问题,又能回答现代人种在非洲汇合杂交的起源。
中国学者应该坦然接受现代人最早起源于非洲说,因为中华民族起源既有土著特点又有非洲特点,而不应该以政治斗争说保护非科学,即认为“非洲起源”说是各种“外来说”的翻版,一方面受着国际流行的学说与方法论的影响,有些也受着西欧中心史观的支配,同时也限于外国的学术水平低与资料缺乏;这些假说提出之后,在中国与西方即已受到了驳难。其实,这个逻辑不成立.可叹中国大多数的传媒记者和专家、学者,陷于政治和脸面而耿耿于怀,一次又一次上演“买椟还珠”的新悲喜剧:把不存在的现代人种起源于中国留给自己、把真实的远古文明起源推给别人。
且不说有陈竺等中国分子生物学家经测定,已证明中国现代各民族的人种和国际科学共同体认定,现代人种起源非洲的基因证据是一致的。而且在四川绵阳涪江和梓江下游区域,距今8000年左右的古山寨聚落遗址犹存,它们规模宏伟,气势壮观;围绕山寨的处于半山腰的大围坪,延伸数百里,境内文物古迹众多。这些与古书记载的蛮荒历史无一相似。这使人想起了20世纪初留学日本归乡的绵阳学人何拔儒,通过长期考察和研究从盐亭辐射嘉陵江流域两岸的大围坪和古山寨地质地貌,得出从1.2万年至8千年前的大冰川末期起,四川曾发生和存在过多次盆塞湖和盆塞海现象,从而为5000年前中国的海洋文明和山寨城邦文明提供了地质地理条件,而被人称为中国上古海洋文明和城邦文明古环境研究的第四部书。
但要解决四川古盆塞海洋文明与中原的古黄土文明谁在先?刘东生院士2002年获得的被誉为诺贝尔环境奖的“泰勒环境奖”,这是认识地球上自然历史、气候、生物变迁的第三本书;即它认为中国的黄土高原已经存在了250万年,我国的干旱史也有250万年,这是投了黄土高原的弃权票。因此,如果说人类最早的顶尖优势文明起源于8000多年前的四川盆塞海洋文明和山寨城邦文明,那么就能说明中国后来的北方、南方以及海外的远古文明渊源问题,因为在这些顶尖优势文明产生之前,嫘祖、夸父、盘古、女娲、伏羲、蚕丛等先王,就已经在四川盆塞内陆海及四周城邦之间演习操练过多时了。
然而在解放前这些真实古史的科学方法的研究,就得不到第一部门(政府)和第二部门(企业)的承认,只能长期埋没于第三部门的极少数如绵阳学人何拔儒这类民间学者的脑中。我们不能不如实报道,1949年新中国的解放,由于革命和政治斗争的需要,四川省仅在盐亭县就有数百特定环境中和革命发展不合拍的人物被镇压;大批收缴中的有关古书籍、古文献资料被焚烧,数十座远古山寨城邦遗存被拆毁。其中,在文革结束前的历次政治运动中,也有消毁证据的事情。今天当不知情者要求拿出第一手远古文明的相关材料时,该不该把这些消毁证据的证据提供给审视者。而今天,60岁以上活着的人中,嫘祖故里四川省盐亭县还能找到成十上百的这些见证人到场作证。
我国古人类学家周国兴说,年代测定早在上世纪80年代,北京大学原思训等人就对柳江人化石地点堆积物的年代进行过测定,除了碳14法外,也采用了铀系法,测得的年代从大于距今4万年、7万年或8万年,直至距今约10万至22万年。而且对柳江人年代的测定数据,均不是来自人骨化石本身,它要么来自地层,要么来自伴生的哺乳动物的骨化石或牙齿化石,故都是间接年代。严格地说,柳江人究竟有多古老,尚难定论。此外,柳江人化石并非正式考古所得,是农场工人挖岩泥时得到的,它的确切出土方位并不明朗,所测数据不一定能准确地反映柳江人的生活年代。正因为此,所以原思训等提供的年代数据并未被古生物学界全部采纳,仅仅作为参考数据,还得参照地层、古生物学、人类化石形态特征以及同期其他古人类遗址等多方面的考量,来考虑柳江人的生存年代,一般认为这一年代在距今3—5万年间。即古生物学界对柳江人年代的认定还存在分岐。但认定它存在于距今7—13万年或更早的研究人员却断定这些年代是具有可靠性的,然而他们所说的“铀系法”,是指“新生碳酸盐岩铀系测年法”。
我国有些学者曾利用此法测定与柳江人地点邻近的柳州白莲洞人遗址,结果与用碳14法所测出的年代相差甚远,竟把白莲洞人牙化石年代从距今3—4万年提前到16万年前,他们还测定了柳江人化石地点的地层,认为柳江人不少于16万年。须知,在这个年代,柳江人生活的溶洞尚未形成,这显然是不符合逻辑的。另外,如果承认柳江人和白莲洞人生活在16万年前,那么它势必成为化石智人的早期代表,但它在形态特征上却与华南地区已知的化石智人早期代表马坝人差距太大,柳江人是现代型,马坝人接近猿人型,这同样是不合逻辑的。综上所述,我们不能认为这种“铀系法”测出的年代一定可靠,任何一种测年法都需要经过多方的验证。
但周国兴也说,有些专家认为,在古文化方面,主要是石器制作技术上承袭了祖先的技术,而看不到来自非洲的因素。这也是对多地区论的支持。我国分子生物学家根据线粒体DNA和Y染色体的研究估算出,现代东亚人群的非洲祖先大约在距今6—1.8万年前首先进入东南亚,再转入中国南方,最后越过长江进入北方地区。这些专家认为中国古人类化石记录中有一个“空白期”,即距今10—5万年间尚未有古人类化石的确切记录。他们将这一现象归于末次冰期降临,使绝大多数生物种类难以存活,人类也因此而灭绝。冰期结束后,来自非洲的现代类型人群进入中国大陆,成了这块土地的新居民。这就是现代中国人祖先“非洲起源论”的来历。这种“非洲起源论”也不是没有破绽的。且不说是否真的存在这个“空白期”,并无充分的科学依据,即使有这个“空白期”,也有些问题不好解释。晚更新世始于距今10—13万年前,最后一次冰期始于距今7万年,最冷的冰盛期是在距今2.5—1.6万年前,非洲来的居民正好赶上最严酷的寒冷环境,如果原来的居民不能生存的话,非洲来的居民又岂能生存下来!
这里周国兴犯了一个不大不小的常识性错误。而排除了这个错误,周国兴的反驳更有力地说明中华民族起源既有土著特点又有非洲特点。这就是第四纪大冰川期和始于距今7万年,最冷的冰盛期是在距今2.5—1.6万年前的最后一次冰期,是两个不同层次和温度差别很大的时期,非洲来的居民在最后一次冰期的寒冷环境能生存,而在第四纪大冰川期我国原来土著特点的居民却不能生存或难于生存。
第七章人类的起源和进化(全部:展开/关闭)
7-1人类的起源
人类起源问题的概述(展开/关闭)| 对人类起源的认识,起初是从比较人与动物关系开始的,主要用的是演绎方法。最早提出“人猿同祖”思想的是具进化思想的先驱者布丰(G.L.Buffon)。达尔文的《物种起源》一书在广泛论述生物进化的基础上断言,“人类的起源和历史也将由此得到许多启示”。此后,赫胥黎通过比较解剖学、发生学、古生物学等研究,证明了人与猿是最近的亲属,并于1963年发表了他的名著《人类在自然界的位置》,明确提出人、猿同祖论。1871年,达尔文又发表了《人类起源和性选择》一书,书中既肯定了人与猿的亲缘关系,如表现在身体结构、心理特征和生理特点方面;又肯定了人与猿在直立、双手、牙齿、脑、智力等方面的区别。达尔文认为,支配人猿分化的不是超自然的东西,而是生物演化的规律,即自然选择(包括性选择)的作用。 当然,对于人类来说,除了生物属性外,还有社会方面的属性。对此,19世纪包括达尔文在内的生物学家都未曾有过令人满意的说明,而马克思、恩格斯在这方面曾作了精辟的论述。 20世纪上半叶,首先是逐步积累起来的化石资料,有力地支持人猿同祖的论述,也为用归纳方法研究人类起源问题奠定了科学基础。如果说在达尔文时代,有许多人相信上帝造人的说教,那末,随着古生物学以及分类学、比较解剖学、生理学、病理学,分子生物学等学科的成就,人类起源于某种似猿的祖先的体质进化过程,已被人们所普遍接受。近几十年来,人类起源的研究深入到社会行为、社会联系、文化发展等各个方面。与它们有关的学科,如社会生物学、人种学、神经生物学、比较心理学都对此作出贡献。 |
| 我们已经知道,人类起源是从攀树的猿、经形成中的人、到完全的人的漫长过程。在这个过程中,体质形态、行为特征、群体关系等方面都有巨大的变化,还出现了原始的文化和意识。具体地说,可表现如下几个方面: 1.直立行走和前肢的解放森林古猿被迫下地后,不得不使整个体质形态向人的方向发展。首先是直立行走。直立行走,使前肢获得进化的机会,并逐渐出现人类特有的稳固下肢。直立行走,也迫使躯干、头骨、牙齿、喉部发生一系列变异,如身体重心下移,脊柱从弓状到曲状(S形),出现颈曲和腰曲。人不是唯一能直立的物种,但只有人站立得最挺直、最自然。树栖的猿类,上肢长于下肢,身体重心较高,脊柱只是微微变成弓形。它们偶尔直立,但很不稳,所以多采用半直立姿势(见图)。 前肢(手)的解放促进脑和感官的发展,从猿到人的过程中,脑量的增加相当迅速。现代的黑猩猩平均脑量为345克(脑重与体重之比是1∶61),人类则增加到1360克(与体重之比是1∶45)。脑的发达又反过来指挥着手。尽管最进步的猿手和最原始的人手在骨节和肌肉的数目以及一般排列上是相似的,但人手在细微结构即韧带和骨骼上都得到特别发展(见图)。 2.工具的使用和制造一般推测,人类的远祖会使用工具,从事最原始的劳动。这个结论来自古生物学的资料,现代高等动物的某些行为也可提供佐证。据报道,西非有人发现黑猩猩曾利用石头砸开油棕果的硬壳;也把棍棒伸进土蜂窝内,然后再从棒上舐食蜂蜜。古道尔还多次看到,黑猩猩能拣起草棍,握紧手掌把叶子捋掉,然后将它插入白蚁巢内钩取白蚁。不过,黑猩猩这样的取食行为是个别的、偶然的,人类不仅在使用、制造工具上比其他动物更有想象力和创造性,而且人类运用工具进行劳动是谋生的主要手段。从这个意义上说,人类的劳动是真正的劳动,这种劳动是从制造工具开始的。对人类祖先来说,粗的树枝可以当作棍棒,地上随手可得的石块也可用来抵抗野兽和打猎。石块比木头坚硬,取其尖利的部分剥去兽皮、切割兽肉,这又是一项新的创造。工具的使用和改进,大大有利于原始人类的发展,因而这种行为方式会不断被选择。 3.脑容量增加和动态持续假说脊椎动物在陆地上的进化期间,从最初的两栖类一直发展到人,脑的相对大小增加了100倍,而且增加的速度也特别快。从猿到人的过程又更为明显(见表7-1)。 脑容量的增加在选择上的意义是容易理解的。因为人类的体力总相当有限,克敌制胜的关键还在于智慧和群体的力量。据哈维兰报道,现代狩猎民族,如南非的桑人(原称布须曼人),在分析猎物的行踪、制定未来的行动方案时,他们头脑的活动就不断加强。古人类如果只依照老一套的本能行为办事,在竞争中是很不利的。只有善于动脑筋的类群,例如能够预测问题、挖坑、设陷阱,或用石头、棍条和骨头制造武器的,才能更好地生存、繁殖和兴盛。 事实上,脑容量的增加和智力进化是呈正相关的。据说(Saqan,1980)在哺乳动物中,唯有人类分娩时才发生阵痛,还常因此伴有难产,这除了因直立行走使骨盆位置变化外还与脑量增加、脑颅增大有关。现代人的脑容量是南猿的2—3倍,是能人的2倍。脑容量增加而导致的分娩阵痛是适应相对性的表现。 人类大脑的进化,不仅表现在胎儿期,也体现在整个个体发育过程中。有人认为,“幼态持续”(neoteny)是实现大脑进化的有效机制。所谓幼态持续是指个体发育中保留某些幼年时期的特征并使之延长到成年时期的现象。有人(如Morris,1969)统计,猴子胎儿的大脑在出生前迅速增大,并趋于复杂。出生时,它的脑容量相当成年猴的70%,余下的30%在出生后6个月内发育完毕。黑猩猩在出生后头12个月内,其大脑才发育成熟。人类出生时的脑容量只相当成年后的23%,出生后的6年内,发育的速度很快,整个过程要到23岁才全部完成。也就是说,对人来说,大脑发育在性成熟10年之后仍在进行。但对黑猩猩来说,在它具有生殖能力前6—7年,大脑就已完成了它的发育。正因为这样,人类大脑才有充分发育、不断完善的机会。 4.食性和繁殖的进步人类远祖的食性兼有灵长类和食肉动物两者的特点,他们既吃植物又吃肉。肉食大都是通过狩猎取得的;它不仅大大增强古人类的体力,更重要的是对脑的发育有很大的价值。恩格斯曾经风趣地说过,请素食主义者先生们原谅,如果不吃肉,人是不会发展到现在这个地步的。此外,食素必须吃大量的植物,需消耗很多时间,像猴类那样不间断地吃着东西。古人类的食性改善后,就改掉了树栖祖先那样的习性,与食肉动物相似,趋向于间断进食。这使他们有更多的机会从事别的活动,有利手和脑的发展。 在所有的灵长类动物中,人类的祖先是性生活最活跃并兼有多种特性的结偶动物。其中最显要的例子是交配方式的改变和受配时间的增加。这方面的进步非常有利于配偶的和睦相处和后代的成长,从而逐步显示出人类性爱的魅力。 结偶和男女分工有利于繁殖的进化。如果狩猎的任务落实到男性身上,那么料理家务、照管子女主要应是女性的事。由于人类的幼仔发育缓慢,对父母的依赖性很大,因此,形成牢固的配偶关系和分工合作的习惯,其进化上的意义是显而易见的。 需要补充的是,上述这些内容主要局限在自然的范围内,如果涉及到社会因素,那就会复杂得多。 5.皮肤的裸露在上一章中曾提到,现存的灵长类动物中,唯独人类除少数几处毛发外,基本上皮肤裸露、浑身光洁。当然,人类的身子不是没有毛,而是说毛已退化,有的也已失去其原有功能。在没有剪刀等工具的情况下,人类的头发任其生长,可超过膝盖。回溯原始时代,我们的祖先光着身体,顶着这样的头发,这副模样与其他猿猴相比,必定是相当醒目和古怪的。有趣的是如在前面说过的那样,人类在胎儿期间,却类似于猿猴,周身覆盖着胎毛。这层毛在出生前就脱落了,真正的“毛人”数量是极少的。 从进化上说,人类这身裸露的皮肤必然有它的特殊价值。不然在自然选择面前是难以通过的。那么,如何解释这种变化呢?有的认为这与降低热量、适应地面狩猎方式有关。在强烈的选择压力下,脱去这身沉重的体毛、增加汗腺至关紧要。同时,又增添了一层皮下脂肪,显然有利于御寒。也许只能是这样才能勉强生活下去。这是一种直接转化的说法。另一种看法是,人类进化过程中可能有过一段水生的时期,无毛是水生哺乳类(如海豚、海豹等)的适应特征。当水栖猿类返回陆地时,嗜食甲壳类动物的习性转为肉食。这是间接转化的说法,也称“海猿”理论。此外,还有一些有趣的假说。由于“海猿”理论没有直接的证据,因此不少人倾向于直接转化的观点。 6.群体关系的发展人类远祖是一种高度社会性的动物,他们有群居生活的社会本能。凡灵长类动物都有这种特征,它们的生活和迁徙活动都是成群结队的。群体中都有自己的首领,每个成员似乎也都知道自己所处的地位。在从猿到人转变过程中,这种社会性的特性得到改善和加强,并成为人类进化的重要因素。首先,人类远祖下地后,在残酷的大自然面前,不得不以群体的联合力量和集体行动来弥补个体自卫能力的不足,并完成狩猎的任务。 其次,早期人类血缘关系上的发展,也是群体关系进步的重要因素。在灵长类的一些动物中,性活动有助于把不同性别的个体组合在一起,从而形成比较稳定的群体。这种具有较大内聚力的群体,自然表现出选择优势。可以想象,在早期人类中也完全可能存在这种情况。然而,雄性优势又往往会破坏群体的巩固和发展。也许由于集体性的活动,如生产上的协作、狩猎等等的发展,某些雄(男)性之间能相互宽容、减少嫉妒,从而出现较大的持久集团。 随着群体关系的进步,最终出现崭新的因素——人类社会。猿的群体关系(与人类出现的同时),演变为原始的人类的社会关系。 7.原始文化和意识的萌芽文化和意识都是人类形成过程中萌发起来的新因素。对早期人类来说,那时的文化主要指语言的创造、最初的生活和生产经验;萌芽状态的意识,就是对个别实际效益及其条件的意识,到后来才发展到对制约着这些效益的自然规律的认识。 文化和意识都属于人类所特有,但任何现存的事物都有它产生和发展的过程。非人灵长类、甚至某些鸟类都具有文化方面的最简单、最基本的特征。我们暂且称它为“动物文化”。在一般情况下,日本猕猴会用前肢擦去甘薯上面的脏物以便食用。有一次,一只小猕猴偶尔将甘薯拿到小溪中去洗,不久,这件事传开了。4年后在这个猕猴群中,居然有80—90%的成员都会用水冲洗甘薯。此外,有些猕猴曾用海水洗过甘薯,也许是盐分改进了食物的味道,其他的猕猴也慢慢学会了这种方法。据古多尔说,有只年幼的雌性黑猩猩,由于失去母猩猩,所以必须照料它的同辈幼体。后来,在它长大后生下第一胎小黑猩猩时,它看管幼仔的能力明显超过别的黑猩猩。出现在猿猴中的这类动物文化,在一些鸟类中也有发现。动物文化可以说是动物劳动或复杂行为所创造的。 意识是人脑的机能和属性,人脑起源于猿脑,似人的猿类和似猿的人类已有较高的智力,这可称为意识的萌芽或动物意识。意识的源泉在于实践,人类远祖的“实践”主要是用手所从事原始劳动。据化石资料判断,南猿能直立行走,表明它已分化出两只手。但脑尚不发达,只稍大于黑猩猩。也就是说,人类在形成过程中,直立行走比发达的脑的出现至少早200万年。到距今约150万年前,才出现有较大脑量的人类。这些事实证明,劳动实践和经验的积累对人类大脑的发展和意识的形成有着决定性的意义。此外,文化和意识的产生除取决于体质形态、原始劳动和群体关系的进步外,还必须借助于语言。语言是思维的工具,也是交流信息的手段。语言的产生是人类的伟大创造。 |
| 1.灵长类的起源现存的灵长类,从低等到高等的排列,有树鼩、狐猴、眼镜猴(即跗猴)、猴(如猕猴)、类人猿(如黑猩猩)和人类。从古生物上研究,一般认为,灵长类最初的代表是树鼩一类的食虫目小动物,猿起源于猴,人起源于猿。大体的过程叙述如下。 灵长类的起源与大陆漂移直接相关。晚白垩纪,随着地球的激烈运动,恐龙绝灭,哺乳动物开始大规模的“适应传布”。由于巨大森林带的扩展,为树栖动物的产生提供了生态条件。树鼩类的小动物大约出现于6700万年前。它们大体以种子、昆虫为食,吻部长而尖,耳小,趾富有柔韧性,适于抓握。它们以抓昆虫为生,必然也相当机敏。从北美和欧洲的一系列化石中获悉,这类动物的脑比树鼩还大一些,化石臼齿与灵长类相似,而大脑的嗅觉神经部位明显小于树鼩的类似区域,而视觉则比较发达。它们的耳朵中等大小,与另一类灵长目动物狐猴相似。 类似狐猴(Lemur spp.)和跗猴(Tarsier spp.)等原猴类(prosimians)动物出现在始新世,约5800—3700万年间。最早的狐猴中有兔猴属(Adapis)和北美的古狐猴(Notharctus),它们与现代狐猴极为相似。 猴和猿类的祖先最早出现在渐新世,约3700万年前。这段时期化石资料很少,大体判断可能是原始的狐猴。旧大陆(主要指亚、欧非洲广大地区)猴出现的第一个证据来自埃及费菜地区渐新世地层中的化石颌骨碎片。它们的齿式虽然原始,但上颌尖的样式很像猴。现代较高等的猴类有两支:南美大陆(属新大陆)的阔鼻猴和非洲大陆(属旧大陆)的狭鼻猴。图7-5上方是两种猴,即鼠猴(squirrelmonkey)和猕猴(macaquemonkey),前者产于新大陆,后者产于旧大陆。有关资料认为,旧大陆猴(也称旧世界猴)与古猿的亲缘关系比较接近。 最早可辨认的猿称原上猿(Dropliopithecus),它的化石也产于费莱地区。它的牙齿和颌骨的深度和现代猿相似,但个体比现代的任何大猿要小得多。原上猿的后代很可能是某些埃及猿(Aegyptopithecus)。从化石资料看,猴和猿类从渐新世起就分化了。类人猿的直接祖先可能就是原上猿,而不是原先说的类似猴一样的动物。关于原上猿的情况,至今还不清楚。 中新世(约2500万年前)的猿类化石很丰富,从长臂猿到大猩猩都有发现。其中的3个属已确认是现代猿的祖先。这些化石猿中,有一种叫上新猿(Pliopithecus),其头骨、齿式与现代的长臂猿相似,它可能是渐新世的爱奥罗猿的后代。那时已绝灭的一种奥里罗猿,它是渐新世的一个属,即apidium的后代。可见,猿类的起源具有多元性。 比上新猿稍大的是森林古猿(Dryopithecus)。在欧亚和非洲都有它们的化石记录。可见,森林古猿分布很广,在灵长类中是很成功的类型。森林古猿的化石有猴类的特征,但它们的下颌具有5个齿尖的臼齿,清楚表明是属于猿类。森林古猿的后代有较大的分化,个体较小的可与现代的俾格米黑猩猩相比,个体较大的则与现代大猩猩差不多。森林古猿属可能是渐新世埃及狼的后代,又是现代猿甚至古人类的共同祖先。 2.西瓦古猿在森林古猿的后代中,有一种称为西瓦古猿(Sivapithecus)的猿类。不久以前,一般认为西瓦古猿与腊玛古猿(Ramapithecus)是两个属,但从80年代以来,在欧洲、巴基斯坦发现的新化石并结合以往的化石资料分析,认为“腊玛古猿根本不是一个单独的属,而是西瓦古猿属的一种较小的变体”,也有人认为腊玛古猿是西瓦古猿的雌性。 西瓦古猿的最早化石年代是中新世中期,大约1500万年前。它们是猿科动物但有一些相似于人科的性状,如牙齿较小、臼齿的釉质较厚、齿弓形状呈V形。特别是在犬齿的位置上面,有些化石呈凹陷状,这种特征除人类外在任何猿类中都未见到(见图)。所以,西瓦古猿在人类起源中的作用,曾引起许多人的兴趣。 从西瓦古猿化石的有关材料分析,它居住的范围很广,包括热带林区和较干燥的灌木区,以及森林和开阔草原的交界处。这样,在森林减少时,它可能从草原上获得种子一类的食物以弥补森林食物的不足;而当发生危险时,森林则为它提供避难场所。因此,与别的猿类相比,在生存斗争中,它们具有相对的优势。此外,西瓦古猿的重心似乎比现代猿低,现代猿偶尔用两足直立,但相当笨拙,而前者两足行走,其姿势就可能不那么别扭。猿类偶尔的直立,也许主要是用前臂挟运食物,它们不像猴子那样可用颊囊运藏食物。直立的行为,自然很冒风险,至少容易暴露。但也使视野宽阔,当食肉兽接近时能及早发现;甚至可以用前肢扔东西,以对付来犯者。有人发现黑猩猩偶尔会拿石头打豹子,那么我们也有理由设想,它们在中新世的前辈,也可能会采用同样的方法。不过,近几年来的研究认为,西瓦古猿更可能是猩猩的祖先。 3.南方古猿南方古猿(Australopithecus)简称南猿。南猿在分类上是人科的一属,即南方古猿属(Australopithecus)。南猿的进步类型能制造工具,是人类形成阶段的代表。 南猿化石最早是1924年达特(R.A.Dart)在南非金伯利附近的汤恩(Taung)发现的。是一块幼儿的头骨化石(见图)。从1936年起,在南非也陆续发现这类化石,标本数量相当多。这类化石可归为两类:南猿非洲种(A.africanus)和南猿粗壮种(A.robustus)。50年代末以来,路易斯·利基(L.S.B.Leakey)首先在东非坦桑尼亚的奥杜韦峡谷,随后又在肯尼亚某些地区也发现南猿化石。最初定名为东非人鲍氏种,后改为南猿鲍氏种(A.boiscei),从而扩大了南猿存在的地区和时间。1974年,约翰逊(D.Johanson)在埃塞俄比亚阿法洼地发现一系列人类化石,其中包括一具女性个体,保存有近一半的化石骨架,取名露西(Lucy)。定名为南猿阿法种(A.afarensis),其时间为350万年。与此同时发现的还有36—65个个体。 一般认为,南猿有两类,一类身体细小、结构也较轻巧的称纤细型,以南猿非洲种和阿法种为代表;另一类身体较重、骨架粗壮、颌骨粗大的称粗壮型,以南猿鲍氏种为代表。 纤细南猿大小和现代的俾格米人相似,估计身高145厘米,体重约27—43千克,脑容量为428—458毫升。大约相当于大型黑猩猩的脑容量,是现代人脑容量的1/3。它们对信息的处理能力大约与现代黑猩猩差不多。纤细南猿的化石遗存表明:早在400万年前,这种古人类已两足直立行走了。不仅它们脊椎骨的弯曲状况与人相像;而且,“露西”的上臂骨比猿类的也短。后者说明上臂比猿类轻,身体重心较低。尽管与人相比,纤细南猿的前臂略长。这表明它的原始性,也可能与一定的树栖行为有关。估计它们在遇到危险时,会跑向附近的树林。从纤细南猿大量的腿骨和髋骨的形状表明,它们能够两足行走(见图)。还有人根据火山灰留下的脚印痕迹分析,它们的形状、两脚间的距离以及“足尖朝外”的程度,都酷似人类。 粗壮南猿比非洲南猿大得多,估计身高为153厘米,体重约36—54千克,脑容量也稍大,约500毫升。但生存年代较晚,最近的看法约为150—200万年。 关于两种南猿类型的关系,主要有两种意见:一种认为纤细南猿是人类的直接祖先,而粗壮南猿则是进化的旁支。据推测,南猿的社会组织十分简单,群体生活过程中可能有简单的语言。纤细南猿是南猿中朝杂食方向发展的一支,它们的食物中肉类成分加大,并已能成功地制造工具。另一种则认为,两者只是不同性别之差,纤细型是雌性,粗壮型是雄性。哈维兰等认为,粗壮南猿实际上具有纤细南猿,尤其是南猿非洲种的所有特征(见图)。 |
| 7-2 人类的进化 |
| 能人(Homohabilis)也称“早期猿人”,他们是早期人类第一阶段的代表。1959,1960年,利基夫妇在坦桑尼亚发现鲍氏“东非人”附近稍低的层位中,发现了儿童的顶骨、下颌骨和手骨化石,以及成人的手骨、锁骨和足骨化石;1963年,又在较高层位里找到一个头骨和下颌骨化石。这些化石类似南猿,在1964年定名为“能人”。能人头骨壁薄,眉脊不明显,平均脑容量为637毫升。颊齿,特别是前臼齿,比南方古猿非洲种为窄。下肢骨明显显示“能人”已能两足直立行走的特征,尽管这些特征远不如现代人那样完善。手骨表明其拇指和其他四指能够对握,但还不很精确。与能人化石同时发现的还有不少石器,典型的石器是用砾石制成的砍砸器,这种石器文化叫做奥杜韦文化(Olduvaiculture)。能人生活层中有被宰杀的动物(包括东非南猿)遗骸,表明他们已经会打猎,也许已能在一个广泛的地区内进行活动。 从60年代后期起,在肯尼亚特卡纳湖的东面和北面发现大量似人化石,其中最好的一个是利基的儿子理查德·利基(R.Leakey)发现的,即著名的KNMER1470号人(KNM代表肯尼亚国立博物馆,ER代表卢道尔夫,“1470”是博物馆标本的收藏编号)。1470号人头骨与任何南猿的头骨相比都要进步,脑容量可达775毫升。肢骨基本上和现代人相似。地层年代原先测为距今270万年,后来的研究结果是200万年左右。这一化石被认为是能人类型的主要代表。近年来,还发现编号为3733号人的头骨7,其年代在130—160万年前。 一般认为,能人是介于南猿和直立人的中间类型。但也有认为,它应属于纤细南猿。 |
| 直立人(Homoerectus)是晚期猿人(也称猿人)的学名。它是早期人类第二阶段的代表。 1891年,荷兰人类学家杜布瓦(F.Dubois)在印度尼西亚爪哇特里尼尔(Trinil)发现了爪哇直立人(H.erectuserectus)。自此以后,在亚、欧、非洲各地都发现类似的化石,地质年代包括早更新世晚期到中更新世。这些直立人之间只有亚种级的差别。后来研究表明,直立人与其前的南猿有属的差别,与其后的智人则是种的差别。 直立人化石中最主要的代表是爪哇直立人和中国直立人(H.erectuspekinensis),但前者比后者更为原始。此外,在中国的还有元谋人、蓝田人(H.erectuslantienensisi),还有印度尼西亚的莫佐克托猿人和直立猿人;非洲有坦桑尼亚的利基猿人、阿尔及利亚和摩洛哥的毛里坦猿人;在欧洲有德意志联邦共和国的海德堡人(Homoerectus heidelber gensis),匈牙利和捷克斯洛伐克近年来也发现了猿人化石。猿人大约生活在200—150万年到20—30万年前的旧石器时代早期。 我国北京周口店的北京人遗址,是世界上至今已发现的材料中最丰富的直立人遗址。经古地磁法测定,北京人的绝对年代不超过69万年,地质年代属更新世中期。第一个头盖骨于1929年12月2日发现。北京人头骨的主要特点是头骨的最宽处在左右耳孔稍上处,而现代人的头骨最宽处则在较高的位置。北京猿人头骨的高度远比现代人为小。额向后倾斜。平均脑容量为1088毫升,而现代人平均为1360毫升。眉脊非常粗壮而前突。头骨的厚度比现代人几乎大一倍。牙齿比现代人的硕大和粗壮。面部比现代人相对地较短而明显前突。北京人大腿骨的主要性状与现代人相似,有股骨脊(股骨后面突出的脊的存在表明已有拉直躯干的发达的肌肉),以及肱骨短于股骨的事实,可以确定北京人已能采取直立行走的姿势,像现代人那样地迈步行走。 我国周口店的北京人文化是亚洲重要的直立人文化。它是以发现诊贵的北京人化石而著称于世的。 北京人石器的主要类型有砍砸器、刮削器和尖状器等几大类。大型的砍砸器多是用砂岩砾石打制的,可以用来砍伐树枝。小型的刮削器,有直刃凸刃、凹刃和多边刃等多种,大部分可能是日常生活的用具。 值得注意的是直立人已开始用火。从北京人遗址中已发现有木炭,很厚的灰烬层,燃烧过的土块、石块、骨头和朴树籽等,表明北京人不仅已经知道用火,还知道保存火种。从伴生动物诸如鹿、绵羊、羚羊、獐等的骨头化石表明,直立人已是熟食的狩猎者。不仅如此,还有食人的风气。魏敦瑞(F.Weidenreich)和贾兰坡等认为,根据周口店混杂在动物化石中人的骨头状况表明,北京人已会吃人。其中某些头骨有被从枕骨大孔打穿进去的痕迹,推测这是古人类为吸取有营养的脑髓而造成的;长长的肢骨的两端被他们劈开来敲取骨髓。 在欧洲,和直立人同时代的石器文化有著名的阿布维利[即舍利文化(Chellean culture)]、阿舍利文化(Acheulianculture)和克拉克当文化。典型的阿布维利文化(Abbrevillianculture)用的代表性的工具是手斧,它是以燧石(chert)结核打制的。阿布维利文化继续向前发展,就进入阿舍利文化,它占优势的工具仍然是手斧。克拉克当文化,是一种以石片为主体的旧石器时代文化。 在直立人阶段,婚姻形式和社会形态已从早先过渡阶段的杂乱性交关系的原始群进入了血缘家庭的前氏族公社。血缘家庭后是群婚,即整个一群男子与整个一群女子互为所有,很少有嫉妒余地的婚姻形式。婚姻集体是按照辈数来划分的。 |
| 化石智人比直立人更接近于现代人,他们是早期人类第三阶段的代表。他们不仅完全直立行走,而且脑量已与现代人相似。因此,他们与现代人属同一个种,即智人种(Homosapiens)。化石智人分为早期智人和晚期智人两类。 早期智人(或古人) 早期智人的代表是尼安德特人 (Homo sapiensneanderthalensis),简称尼人。“尼安德特”是德语中德国杜塞尔多夫附近一个山谷的名称。1856年,即达尔文《物种起源》出版的前3年,在尼安德特山谷一个斜坡的洞穴中发现了这个早期人类。 本世纪50年代末,在亚、非、欧洲广大地区己找到大量的早期智人化石。其中有人骨化石130余件,遗址50余处。我国在广东、湖北、山西等省都有发现,主要有马坝人、长阳人和丁村人。早期智人已有与现代人更接近的特征,但仍带有相当多的原始性质。他们大约生存于距今2、3万年到4—5万年前,已进入到旧石器时代的中期。在文化上,他们已能制作几种式样不同的标准化的石器。同时不但能使用天然火,而且能人工取火了。 早期智人化石除美洲、大洋洲外,各个大陆上都有发现,数量很多。地层年代介于直立人和晚期智人之间。这些化石差异较大,有的时代晚反而形态较原始。因此对这些化石之间的关系以及在人类发展史上的位置有多种说法。比较普遍的说法是认为发现于西欧大陆的许多时代较晚的化石属于典型尼人,其他的称为非典型尼人。在非典型尼人中有的形态较近典型尼人,被称前尼人。有的形态较近晚期智人,则称前智人。 典型尼人的化石经复原后,可见眉脊发达,头顶低平,额部很倾斜但不缩狭,头骨最宽处比现代人低,但比猿人的稍高。头骨后部仿佛被从上下方向压挤了似的,不像现代人那样圆隆,颞骨鳞部的上缘几乎成一直线,不像现代人那样成一向上凸的弧线,这些特点都反映了脑子外形较为原始。这一典型尼人脑于的体积较大,脑量定为1575毫升。脑子比晚期猿人大,脑颅的形状介于直立人和现代人之间。尼安德特人也属于典型尼人。 早期智人的脑量已达到现代人的水平,远比猿人的脑大和复杂,已经有比猿人更高的智慧。他们制作的石器有了很多改进,人们推广了一种打制石片的新技术——修理石核技术。在西欧,以手斧为主体的阿舍利文化逐渐被以细小尖状器和刮削器为代表的莫斯特文化(Mousterianculture)所代替。早期智人能够狩猎巨大的野兽,并能用兽皮当作粗陋的衣服,不仅会使用天然火,也可能已会取火,因而能适应各种气候条件,他们分布于亚、非、欧三洲的广大地区,生活了很长的时间。不仅如此,尼人开始有埋葬死者的习俗,而且也已穿着衣服,不再像猿人那样赤身露体了。 尼人阶段的人类社会形态,已从原始公社前一阶段的前氏族公社进一步发展成原始公社后一阶段的氏族公社(母系氏族社会)。这时的婚姻关系从族内婚进到族外婚,也就是一氏族的成年男子集体到另一氏族,与该氏族的成年女子结为婚姻,但死后仍安葬在本氏族的领地内。 晚期智人(或新人)晚期智人在欧洲的代表是克罗马农人(Cro-MagnonMan)。他们是早期智人的后裔。克罗马农人的化石最早是1868年在法国南部克罗马农洞的裂隙中发现的。 早期智人是怎样进化到晚期智人的?有一些不同的看法。布雷斯(L.C.Brace,1964)认为,后期的尼人到早期的克罗马农人之间有5千到1万年之久。选择的压力足以使他们实现这一转化。这些压力主要是文化上的,并且与使用工具(如刀)和食肉有关。 克罗马农人生活在旧石器时代的晚期,约距今3.5万年前。他们生活的时期比尼人要短得多。但他们的化石分布却比以往任何阶段的人类广得多,数量也多得多。不仅在欧洲、亚洲和非洲,大洋洲和美洲也有他们的遗迹。从1872年到1902年期间,在意大利的里维耳拉附近就出土了13个化石标本。在威尔士发现的一具无头的骨骼也被纳入克罗马农“家族”。在我国的晚期智人有华南的柳江人、四川的资阳人、内蒙的河套人和北京周口店龙骨山的山顶洞人等。 晚期智人与早期智人相比,在体质特征等方面有明显的进步。以克罗马农人为例。他身材高大,长达180厘米。颅骨的高度增大,额部隆起,有明显突出的下颏。他们四肢的特点是前臂比上臂长,小腿比大腿长。脑子已很发达,有相当的智慧。 欧洲晚期智人的文化分为奥瑞纳(Aurignacian)、梭鲁特(Solutrian)和马格德林(Magdalenian)三期。晚期智人更多地以石叶来制作工具,石叶也是一种石片,但比石片要窄长。他们除了石器比古人加工精细外,还有不少骨器和角器。晚期智人不仅能使用简单的工具,还能用石头或骨角制成矛头,加在木棒上,制成长矛或标枪等复合工具。他们的狩猎能力有了进一步的发展,已知利用地形狩猎,甚至会用陷阱捕捉大兽。他们已有了相当好的捕鱼技术,还会用大兽骨、鹿角、象牙、兽皮等修建成简单的房屋。而且还能够摩擦生火。 晚期智人阶段出现了原始的艺术,在他们的洞穴里发现过不少壁画。如西班牙阿尔塔美拉(Altamira)洞中的野牛、野马、鹿及其它动物的画像,笔法熟练,有浮雕感,可算是洞穴艺术的代表作。原始艺术品内容大都与狩猎活动有关,这表明艺术从一开始就是与生产实践有着密切关系的。在晚期智人的文化遗物中,还有不少装饰品。裴文中详细研究了山顶洞人的文化遗物,他发现其中有穿孔的兽牙、穿孔的海蚶子壳、钻孔石珠等等,并以穿孔兽牙为最多。贾兰坡指出,原始人类从事装饰的目的,不仅是为了显示他们的英雄和智慧,而且与吸引异性有关。 晚期智人的文化,除旧石器时代外,还包括中石器和新石器时代。中石器时代的文化特征,是广泛地使用弓箭以及镶嵌工具,由于弓箭和其他复合工具的制造和使用,促进了生产的提高,因而在经济上也起了变化,即由采集和狩猎的掠夺性经济向耕种和放牧这类生产性经济过渡。在某些地区,人们还驯养了狗。而新石器时代则出现了原始的手工业(如陶器器皿)和纺织业(如陶纺轮)。 当时的社会,男女已有明确分工。男人打猎捕鱼,女人采集、管理氏族内务。由于男人打猎捕鱼,收获没有保证,而女人采集植物性食物,收获较为稳定,因而妇女在社会生活中占有显著地位。当时的人只知其母,不知其父,妇女是氏族的中心。后来,当畜牧业和农业成了氏族公社生产的主要内容后,男人的劳动在生产中占了主要地位,于是氏族的首领逐渐由男人来担任,父系氏族代替了母系氏族。氏族公社实行原始公有制,共同生产,共同分配,没有人压迫人的现象。不过,这种制度是以生产极不发达为前提的。因而广大地区内人口极度稀少;因此,人类差不多完全受着陌生的、对立的、不可理解的外部大自然的支配。 在晚期智人阶段,现代人种开始分化和形成,分布到世界各地。当然,晚期智人也可叫现代人,但现代人的概念,局限于新石器时代以后的人类,也就是公元前1万年到现在的人类比较合适。 |
| 人种也称种族。它是指体质形态上具有某些共同遗传特征(如肤色、发色、发形、眼色、血型等)的人群。总的来说,不同人种中的某些基因的出现率是有所区别的。例如,某一血型(如O型)在某一人种中可以很高,而在另一人种中则很低。各个人种之间能广泛地进行基因交流,任何人种中都不存在独有的特殊基因。这充分说明全世界各人种在生物学上同属一个物种,并具有共同的祖先。大多数人类学家把全世界的人分为三个或四个主要的人种。这就是黄种或蒙古利亚人种(Mongoloid)、白种或欧罗巴人种(Europoid)、黑种或尼格罗人种(Negroid)。如分成四个人种,则除上述三个外,还有棕色或澳大利亚人种(Australoid)。 黄种主要分布在亚洲和北美洲北部和南美洲北部的部分地区,因而又叫亚美人种;白种主要分布在欧洲及亚洲、美洲、北非和大洋洲的部分地区,因而又叫欧亚人种;黑种主要分布在非洲的赤道附近地区,因而也叫赤道人种;棕种主要分布在大洋洲。四个大的人种又各分为许多小的人种。 黄种人的主要特征是肤色淡黄、脸扁平、鼻中等宽、发形直而较硬;白种人皮肤白、鼻子狭窄、眼睛颜色和头发类型多种多样;黑种人皮肤黑、嘴唇宽厚、鼻低而宽、头发卷曲;棕种人皮肤黄褐或棕黑色、鼻短而宽,唇较厚、头发波形或卷曲。 不同人种所具有的那些共同特征是如何形成的呢?库恩(C.Coon)等人在《种族》(1950年版)一书中指出,主要是由于对环境(主要是气候)的适应而造成的。而且随着诸如基因流动和选择力的改变,人种也在不断地变化。 人种(或种族)与民族的涵义不同。民族不是由体质特征来区分的,而是由具有共同的语言、地域、经济生活等特征来区分的。它是在历史上组成的人们共同体。一个种族可以包含一个或几个民族,例如黄种包括东亚许多国家的许多民族。同样,一个民族也可包括一个或几个种族。例如,我国是一个多民族的国家,由50多个兄弟民族组成,主要是黄种人,但也混有白种人的血统。因此,人种(或种族)和民族是两个不同的概念。 |
| 根据现有化石的推断,现代的主要人种形成的时期,大约在晚更新世的旧石器时代晚期甚至中期开始,距今至少有4至5万年之久,到1万多年以前旧石器时代晚期之末,各主要人种已具有明显的特征。例如,柳江人和山顶洞人即是原始黄色人种代表,是现代中国人的直接祖先。在他们身上已有体质南北分型的倾向。 关于人种形成过程,化石资料极少,在此仅作简要介绍。 黄种(蒙古人种)黄种人分布于亚洲大陆东半部、太平洋群岛以及南北美洲大陆,包括现代的中国人、蒙古人、日本人、印第安人等等。黄种人约在旧石器时代的晚期,起源于亚洲中部和东部。。日本国土当今虽四周皆海,但在第四纪更新世时,它与大陆广泛相连。大陆的动物和人类源源不断迁徙过去。所以日本人同样起源于东亚大陆。此外,黄种人也向欧洲和澳大利亚扩展。例如,在亚洲北部与欧洲人种相混杂,形成了乌拉尔人种和南西伯利亚人种;在亚洲南部与澳大利亚人种相混杂,波利尼西亚人种可能主要是这种混杂的产物。 部分黄种人还可能由亚洲东北部经由最后冰期中干涸了的白令海峡(BeringStrait)进入美洲。较早去的成为印第安人,最晚去的成为爱斯基摩人(Eskimo),后者居住在北冰洋沿岸地区。 白种(欧罗巴人种)白种人分布于欧洲、北非、西亚和北印度等地。近几百年来,随着殖民主义的扩张,又逐渐分布到美洲、南非和大洋洲等地。约在旧石器时代的晚期,白种的基本人群起初在地中海沿岸,即南欧、北非和西南亚的某些自然分布区。后来逐渐散居到整个欧洲和北非。同时,有一部分不断向外扩展。例如,其中有的支系与其他人种,特别是黑人长期混杂,形成了过渡性的类型,如东非的埃塞俄比亚人种;其中的南欧人种,大约是公元前4千至3千年前扩展到印度南部,与古代维达人混合而形成南印度人种。 欧洲的其他人种,例如波罗的海人、诺的克人等,都是由南欧逐渐向北迁徙过程中分化而成的。如北欧人种,由于寒冷与多云的气候条件而形成了浅淡的肤色、发色和眼色。 黑种(尼格罗人种)黑种人分布于非洲、大洋洲、印度南部、斯里兰卡、美拉尼西亚、加里曼丹等地。约在旧石器时代晚期在南亚首先形成非洲黑种人的祖先类型,然后向西南方迁徙,途经阿拉伯半岛进入非洲东北部。然后再扩大到非洲的其他地区。 棕种(澳大利亚人种)棕种人分布于澳大利亚、新西兰、美拉尼西亚、波利尼西亚、新几内亚、斯里兰卡、印度南部等地。可能是旧石器时代中期起源于南亚大陆,向东南方的大洋洲迁徙,从印度支那经粪他群岛到达澳大利亚。澳大利亚与亚洲大陆隔离甚远,澳大利亚人种是在长期隔离的情况下形成的。 |
| 系统说和迁徙说这是有关人种起源的两种学说。 第一,系统说,也称直接演化说,系统说认为,各地区的人种是土生土长的,是由当地的直立人亚种逐渐演化而成,所以系统说也叫多中心说。它认为,不可能在几万年内由一个地区向外扩展而形成现在的各个人种。但不同直立人亚种怎样获得共同的现代智人的特征(如发达的颏突、隆起的额部、充足的脑量)则难以解释。主张这种观点的学者主要有魏敦瑞(1946)、库恩(1962、1965)、布罗斯和沃尔波夫(Brese、Wolpoff,1971)。 第二,迁徙说,也称代替说。它认为各人种是由一个已具有现代人形态的人群迁入、代替(消灭)当地原有的人群而形成的。迁徙说也称单中心说。但这个现代人的起源中心还不能肯定。过去认为在西亚;近年来在南非也发现了较早的现代人化石,因而有人主张南非是现代人种的起源地。主张这种观点的有罗金斯基、埃德沃兹(A.Edwards)等。 近年来,东亚、非洲、欧洲和其他地区,智人化石的发现不断增多,对人种的分子人类学研究也日益深入,从而为尽快解决人种起源问题提供重要条件。 人种理论与种族主义种族主义是资产阶级在人种问题上宣扬种族歧视的一种伪科学。白人种族主义者人为地区分为所谓“优等”和“劣等”人种,从而歧视、迫害和不平等地对待有色人种,尤其是黑色人种。他们片面夸大各人种之间智力和体质上的差异,用来作为制定优等种族与劣等种族的根据。 其实,种族歧视是一个政治问题、阶级关系问题;人种问题的研究则是一个学术问题,两者之间没有必然的联系。它们是两回事,不是一回事。即使各人种之间确实存在着智力上或体质上的某些较大差异,也决不能作为种族歧视的一个理由。这是首先要讲明的问题。况且,实际情况也完全不是像种族主义者所说的那样。上面已经说到,现有的一切民族,都属于人类发展史上的同一个阶段,属于同一个物种。而且,各人种之间,有各种中间类型的存在。不同种族类型接触的地区总是种族混杂的地带。 根据人种特征总的进化过程,的确可以说某一人种的某种特征是比较原始的或是比较进步的。可是在每一个人种中,都同时存在着比较原始和比较进步的特征。以体毛的多少为例,在人类进化过程中体毛是逐渐减少的,而在三大主要人种中,黄种人的体毛最少,白种人的体毛最多;以面部突出的程度为例,在进化过程中,突出的程度是逐渐减小的,而现今是白种人最小,黄种人最大;再以嘴唇的厚度为例,前面已经讲过,在进化上应在黑色人种占优势。 种族主义者还鼓吹所谓北欧种族是人类最有才能的种族,也是毫无根据的。诚然,那里的科学技术与生产比较发达,平均生活水平也比较高,但是在几千年前当波罗的海沿岸的北欧人都还处在文化黑暗的时期,人类的创造智慧已经在中国、印度和孟加拉国的恒河流域、古埃及和美索不达米亚等地取得了辉煌的成果。中国人、古埃及人都不是白色人种。可见种族主义者这类论调也是不值一驳的。 推行种族歧视并妄图达到某种罪恶目的的人,无论过去或现在都不乏其人。希特勒纳粹分子打着雅利安人是优等种族的旗号、疯狂残杀几百万犹太人就是典型的例子。其实,所谓的“雅利安种族”只是个语言上的共同体而根本不是一个种族;“犹太人种族”也只是一个民族类别或宗教类别,更不存在什么种族“低劣”的问题。 人种形成的主要因素人种形成是一个很复杂的过程,往往受到多种因素的制约,其中起决定性作用的是生产,以及与此密切有关的选择、隔离、迁徙、人种混杂等等。例如: 生产:人类社会的生产(包括工具的生产和人类自身的生产)和受其制约的社会制度、文化水平等对人类体质的形成起着主导的作用。例如,人类不是直接地与自然环境发生联系,而是通过物质生产的手段而与自然环境相联系。生产的发展迅速地改变着人类的生存条件。例如,迄今为止,人类的配偶选择在很大程度上与生产水平和血缘关系相关,不完全受性选择所制约。再如,迁徙的发生与生产、血缘关系、经济条件、交通等也是不可分的。在农业开始以前的整个人类历史上,人群经常处于迁徙之中。因此,人类生存条件、婚姻关系、生态环境等都直接影响人种的形成。正如恩格斯在《家庭、私有制和国家的起源》一书中所指出:“根据唯物主义观点,历史中的决定性因素,归根结蒂是直接生活的生产和再生产。但是,生产本身又有两种。一方面是生活资料即食物、衣服、住房以及为此所必需的工具的生产;另一方面是人类自身的生产,即种的蕃衍。一定历史时代和一定地区内的人们生活于其下的社会制度,受着两种生产的制约:一方面受劳动的发展阶段的制约,另一方面受家庭的发展阶段的制约”。 选择:人种的肤色、发形等与选择的作用关系极大。长期生活在横跨赤道的非洲和西太平洋赤道附近的黑人,具有深黑的肤色,就与此有关。皮肤的黑色是黑色素多的原因,黑色素有吸收太阳光中紫外线的能力,从而有保护皮肤内重要结构的作用,可以避免被过多的紫外线照射而受到的损害。黑人卷曲的头发,是抵抗强烈阳光的一种优良的不导热的绝热体。它在头顶形成一个多孔隙的覆盖物,类似毡帽的作用,在赤道的炽烈阳光照射下,由于卷发的导热性差,而不会把外表的热度大量传往头部的皮肤和血管,因而起到了一定程度的隔热作用。 黑人宽阔的口裂,很厚的嘴唇以及短而粗的鼻腔都是自然选择的产物。它增加了水分的散失面积,有助于对吸入空气的冷却,有利于在炎热气候的条件之下生活。相反,在寒冷地区生活的白人,鼻子是高高的。鼻腔粘膜面积明显增大,冷空气吸进时可在鼻腔内加温,起到保护肺部的作用。 隔离:人种的分化与隔离作用有很大的关系。如高山、深谷、河流、沙漠等天然的障碍,使这一区域的人和另一区域的人断绝了往来,长时期在不同的自然条件下生活,发生了种族上的分化。 迁徙:人种的形成与迁徙也是分不开的,许多人种在形成过程中都有过迁徙的历史。迁徙有利于扩大和改善人类的生存条件,有利于不同人种的交往和混杂。 人种混杂:人种混杂也是影响种族发展的重要因素之一。这一现象很早就已发生。而且其规模日益扩大。现在已几乎扩展到整个世界,上述许多中间类型的人种都是长期混杂的结果。随着人类物质文化的进步和交通的发达,各人种间的混杂将更加频繁,其界限将最终消失,当然,这是极其遥远的事情。 以上多种因素又往往是相互联系,相互影响的。许多人种在历史上曾经多次往复迁徙,又经过人种的混杂过程。混杂产生的人种类型,以后又可能长期处于隔离状态,受到新环境的影响而产生新的类型。同时也不是所有人种的体质特征都可以简单地用自然条件来解释的。例如非洲有些黑人现在住在密林的树荫下面,阳光照射不多,但皮肤仍是很黑。其原因可能是其祖先原来生活的是另一种环境,不久以前才搬进森林的。又如黄种人特征之一的眼皮的蒙古褶(Mongolianfold),又称内毗皱襞,在爱斯基摩人中的百分率不高,在美洲的印第安人中也很少出现。黄种人的这种明显的特征,可能是在比较晚的时期生活于空旷的干旱风沙地带的适应性结构。 |
| 7-3 人类的未来 |
| 人类产生到现在这段历史时期,相对于整个宇宙来说,仅仅是其中的一瞬间;这段时间即使与生物进化史相比,也是十分短暂的。为了说明这个问题,我们可以把宇宙大爆炸后的150亿年时间压缩成一年,称为宇宙年。在这一宇宙年内,9月初地球还没从星际物质中形成,恐龙仅出现在圣诞节前夜,开花植物首次出现在12月28日,人类是在除夕晚上10时半诞生的。到了11时46分,北京人开始用火。所有人类有记载的历史都集中在12月31日的最后10秒钟内。例如,指南针的发明在晚上11时59分53秒;欧几里得几何学诞生于59分56秒;以后的3秒钟,就是举世闻名的欧洲文艺复兴。假定把从生命起源到今天这段时期压缩为一天,即24小时。我们也可对此制定一张生物进化的日历表(见图)。根据表的推算,人类最初出现就在晚间11时59分,人类的全部历史记录不过占一天中的最后一秒。 我们作上述这样的比较是要说明,人类的历史是十分年轻的。尽管与别的生物相比,人类的诞生在进化史上占有至高无尚的地位,但人类毕竟还处在幼小的年代。她一定有,也应当有更加光辉灿烂的未来。为了达到这个目的,在很大程度上,将取决于社会进化的水平。 从进化论的角度讲,人类进化是生物进化的高级阶段,它的主要内容是指自然选择作用推动下人类体质和行为特征的进化。但当人类诞生、有了社会以后,使开始了社会进化的历程。社会进化是生物进化的继续和发展,从化学进化到生物进化再到社会进化,构成了宇宙的系统发展。在社会进化过程中,自然选择的作用将受到社会选择的制约,但人类的自然进化仍在继续。例如,某些正在进行中的变化,如“头发、智齿(wisdomteeth)、小趾和阑尾可能逐渐消失”(S.L.Weinberg,1974),人种的界线逐渐缩小等。在人类进化中,比较主要的两个方面是优生与健康。 |
| 7-4 人类起源动力的探讨 |
| 人类起源的动力是一个理论问题,也是个哲学问题。对此,恩格斯早在《劳动在从猿到人转变过程中的作用》(1876年)这篇著名哲学论文中作了精辟的论述。其后100多年来,许多人经过研究,又提出了不少很有价值的见解。 |
何新:中国之名的考古起源
人类的起源及其智力的发展
第十三章 人类的起源与进化
非洲,还是四大洲--人类起源再起争议
【转载】人类的起源与华夏种族的形成【孔子归来】博客:人种:迁徙与和谐【转载】现代中国人的远祖从哪里来?
帝喾祝融及中国人种起源_武陵君_新浪博客
【转】灭绝小黑人创造英国巨石阵和古文明?【转】不肖国人数典忘祖:巨石文化起源于中国论中华文明起源及其早期发展的基本特点(陈连开)李安山:世界历史进程中的非洲-中国社会科学网李安山:世界历史进程中的非洲(1)_文史哲教 光明网人类的起源与中华民族的形成(一)薩尼婭的博客
人类的起源与中华民族的形成(二)薩尼婭的博客人类的起源与中华民族的形成(三)薩尼婭的博客人类的起源与中华民族的形成(四)薩尼婭的博客