裸子植物的孢子体发达,占绝对优势。多数种类为常绿乔木,有长枝和短枝之分;维管系统发达,网状中柱,无限外韧维管束,有形成层和次生结构。除买麻藤纲植物以外,木质部中只有管胞而无导管和纤维。韧皮部中有筛胞而无筛管和伴胞。叶针形、条形、披针形、鳞形,极少数呈带状;叶表面有较厚的角质层,气孔呈带状分布。
配子体退化,寄生在孢子体上,不能独立生活。成熟的雄配子体(花粉粒)具有4个细胞,包括1个生殖细胞、1个管细胞和2个退化的原叶细胞。多数种类仍有颈卵器结构,但简化成含1个卵的2~4个细胞。
裸子植物的胚珠和种子裸露。裸子植物的雌、雄性生殖结构(大、小孢子叶)分别聚生成单性的大、小孢子叶球,同株或异株;大孢子叶平展,腹面着生裸露的倒生胚珠,形成裸露的种子。种子的出现使胚受到保护以及保障供给胚发育和新的孢子体生长初期所需要的营养物质,可使植物度过不利环境和适应新的环境。小孢子叶背部丛生小孢子囊,孢子囊中的小孢子或花粉粒单沟型、有气囊,可发育成雄配子体,产生花粉管,将精子送到卵,摆脱了水对受精作用的限制,更适应陆地生活。少数种类如苏铁属(Cycas)和银杏(Ginkgobiloba),仍有多数鞭毛可游动。由此可以说明,裸子植物是一群介于蕨类植物与被子植物之间的维管植物。
花粉成熟后,借风力传播到胚珠的珠孔处,并萌发产生花粉管,花粉管中的生殖细胞分裂成2个精子,其中1个精子与成熟的卵受精,受精卵发育成具有胚芽、胚根、胚轴和子叶的胚。原雌配子体的一部分则发育成胚乳,单层珠被发育成种皮,形成成熟的种子。
裸子植物常具多胚现象,多胚现象(phlyembryony)的产生有两个途径:一是简单多胚现象(simplepolyembryony),由一个雌配子体上的几个颈卵器同时受精,形成多胚;另一是裂生多胚现象(cleavagepolyembryony),仅一个卵受精,但在发育过程中,原胚分裂成几个胚。一、苏铁纲(Cycadopsida)
(一)基本特征
常绿林植物,茎干不分枝。羽状复叶。大、小孢子叶异株。精子具有鞭毛。
(二)代表属种
苏铁为雌雄异株,大小孢子叶球均集生茎顶。小孢子叶球长椭圆形,由鳞片状的小孢子叶螺旋状排列而成,每个小孢子叶的背面生有许多由3~5个小孢子囊组成的小孢子囊群。大孢子叶球球形,大孢子叶密被淡黄色绒毛,上部羽状分裂,下部为狭长的柄,柄的两侧生有2~6枚胚珠。一层珠被,珠心顶端有储粉室(pollenchamber),珠心中的大孢子母细胞经减数分裂形成胚囊(大孢子),进而胚囊形成2~5个颈卵器,成熟的颈卵器有1个卵细胞和2个颈细胞。小孢子叶上有小孢子囊(厚囊型发育),小孢子母细胞经减数分裂,形成小孢子。成熟的小孢子,进入花粉室,生出花粉管,在花粉管中形成2个陀螺形、有多数鞭毛的精子(图13-22)。精卵结合后形成合子,胚珠发育成红色种子(种皮三层,胚双子叶、胚乳丰富),继而长成为新的植物。
二、银杏纲(Ginkgopsida)
(一)基本特征
本纲特征同属种。
(二)代表属种
银杏(Ginkgo bilobaL.),中生代孑遗的稀有用材树种,又称为白果、公孙树,为落叶乔木,树干高大,枝分顶生营养性长枝和侧生生殖性短枝。年轮明显。各种器官内均有分泌腔。叶扇形,有柄,长枝上的叶大都是先端2裂,短枝上的叶常具波状缺刻,具分叉的脉序,在长枝上螺旋状散生,在短枝上簇生。
球花单性、异株。小孢子叶球呈柔荑花序状,生于短枝顶端的鳞片腋内。小孢子叶有1短柄,柄端常有由2个小孢子囊组成的悬垂的小孢子囊群,精子具多数鞭毛。大孢子叶球简单,通常具长柄,柄端有2个环形的大孢子叶,称为珠领(collar),也叫珠座,大孢子叶上各生1~2个直生胚珠,但通常只有1个成熟,种子近球形,核果状,熟时黄色,外被白粉,种皮分化为3层:外种皮厚,肉质,并含有油脂及芳香物质;中种皮白色,骨质,具2条纵脊;内种皮红色,膜质。胚乳丰富肉质。胚具2枚子叶,不出土,染色体:n=12。(图13-33)。
三、松柏纲(球果纲)(Coniferopsida)
(一)基本特征
常绿或落叶乔木、稀为灌木,茎多分枝,多数种类有长、短枝之分,具树脂道(resinduct)。叶单生或成束,针形、鳞形、钻形、条形或刺形,螺旋着生或交互对生或轮生,叶的表皮常具较厚的角质层及下陷的气孔。孢子叶球单性,同株或异株,孢子叶常排列成球果状。小孢子叶球单生或组成花序,由多数小孢子叶组成,每个小孢子叶通常具2~9个小孢子囊,精子无鞭毛。大孢子叶球球果状,由三至多数大孢子叶(珠鳞,ovuliferousscale)和苞鳞组成,大孢子叶内侧生1~2枚倒生胚珠。种子核果状,胚具子叶2~18枚,胚乳丰富。松柏纲的植物的叶多为针形,故有针叶树或针叶植物(conifer)之称。
(二)代表属种
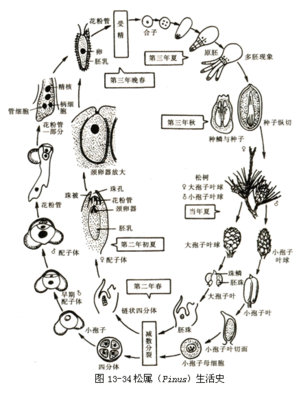
(1)松属(Pinus)
本属植物为常绿乔木,叶针形,常2、3、5针一束,生于短枝顶端,基部全为膜质鳞片。雌(大孢子叶球)、雄(小孢子叶球)同株(图13-34)。
小孢子叶球(又称雄球花,由若干小孢子叶组成),生于当年新枝基部的鳞片叶腋内,多数密集。每一小孢子叶球有1个纵轴,轴上螺旋排列着小孢子叶。小孢子叶背面(远轴面)有一对小孢子囊(花粉囊),小孢子囊中有多个小孢子母细胞,经减数分裂各形成4个小孢子(单核花粉粒)。小孢子经3次不等分裂,形成具4个细胞的花粉粒(雄配子体)。成熟的小孢子具有2个退化的原叶细胞(prothallial)、1个大的管细胞(tubecell)和1个较小的生殖细胞(generativecell),外壁向两侧突出形成气囊。小孢子囊破裂后,花粉粒散出,随风飘扬。
传粉后,小孢子经珠孔进入珠粉室,并萌形成花粉管。在花粉管内,生殖细胞分裂形成2个不具鞭毛的精子,精子随花粉管进入颈卵器,并与卵受精发育成胚。胚珠发育成种子,大孢子叶球发育成松球果。种子成熟后,珠鳞张开,种子散出。种子在适宜的条件下萌发,形成新的植株。
松属中常见的种类有:马尾松(P.massoniana),2叶一束,长而柔;油松(P.tabulaeformis),2叶一束,较短而硬;白皮松(P.bungeana),3叶一束,树皮灰白色;红松(P.koraiensis),5叶一束。
四、买麻藤纲(Gnetopsida)
(一)基本特征
次生木质部具导管,无树脂道。单叶对生,鳞片状或阔叶。孢子叶球序二叉分支,孢子叶球有类似于花被的盖被,或有两性的痕迹。胚珠具一、二层珠被,有珠孔管(micropylartube)。精子无鞭毛,颈卵器极其退化或无。成熟大孢子叶球球果状、浆果状或细长穗状,种子由盖被(假花被)发育而成的假种皮包裹,种皮1—2层,胚乳丰富,胚具二子叶。为裸子植物与被子植物的过渡类群。
(二)代表属种
(1)麻黄属(Ephedra)
植株为多分枝的小灌木。枝条绿色、多节;叶通常退化成膜鞘,对生或轮生,外形很似木贼;雌、雄球花异株;种子成熟时花被为肉质,色鲜红,包于种子之外。如草麻黄(E.SinicaStapf.)是我国西北与华北常见的药材(图13-38)。
(2)买麻藤属(Gnetum)
木质攀缘而缠绕的藤本植物。茎筒形,节部膨大;叶全缘对生,羽状脉。雌配子体无颈卵器。常见有买麻藤(Gnetummontanum Markgr.)和小叶买麻藤[G. parrifolium(Warb)G.Y.Cheng],药用能祛风、行血、消肿、止痛,生汁内服解蛇毒(图13-6-7)。
仅百岁兰[W. bainesii (HK. F.)Garr.]1种,典型旱生植物,分布非洲西南沙漠地带。茎短而粗、块状,终生只有一对大型带状叶片,叶长2~3米,可生存百年以上。雌配子体无颈卵器(图13-40)。
五、裸子植物的经济价值 裸子植物是林业生产上的主要用材树种,我国用在建筑、枕木、造船、家具上的大量木材,大部分是松柏类植物,如东北的红松,南方的杉木。裸子植物又是重要的工业原料植物,可以提供松节油、松香、单宁、树脂、栲胶等,在人民生活中都有重要的用途。
大多数裸子植物为常绿树,树冠美丽,在美化庭院、绿化环境上有很大价值。世界五大庭园植物雪松、南洋杉、金钱松、日本金松和巨杉都是裸子植物。我国的黄山松、水杉、水松、侧柏、龙柏等作为园林观赏树种,也为人类带来了美的享受。
有些裸子植物的种子,如银杏、华山松、红松、香榧等的种子可供食用,如松子等。或作药用,如红豆杉属植物全株含三尖杉生物碱,供作抗癌药物;草麻黄为著名中药材,含麻黄碱,枝叶有镇咳、发汗、止喘、利尿等功效,根可止汗。
我国由于特殊的自然环境——第四纪冰河分散,使裸子植物在我国的残余种类特别丰富,很多稀有种类生活到现在,如我国特产的银杏、金钱松、水松、水杉、银杉、白豆杉等,都是地史上遗留的古老植物,因而叫做“活化石”,上述6种植物也是我国特有的单种属植物。裸子植物中的珍稀濒危植物很多,它们在研究地史和植物界演化上具有极其重要意义,但由于天然更新能力弱而造成资源日益枯竭,目前这些濒危植物种的数量已很少。因此,必须采取有效措施,杜绝乱砍滥伐,保护好现存树种。同时,还应开展繁殖技术的研究,在适宜地区大力育苗造林,以求缓解珍稀濒危植物的生存危机。六、裸子植物的起源与演化在石炭纪、二叠纪的地球植被中,除了外貌像蕨的种子蕨之外,还有一类高大乔木状的种子植物——苛得荻,其植物体为高大乔木,茎粗一般不超过1m,茎干的内部构造和种子蕨颇相似,但木材较发达而致密,木质部或薄或厚,通常无年轮,髓由许多薄壁细胞横裂成片组成,似被子植物胡桃(Juglansregia)的髓;具较发达的根系和高大的树冠,叶皆是全缘的单叶,形态大、小颇不一致,其上有许多粗细相等、分叉的、几乎是平行的叶脉;大、小孢子叶球分别组成松散的孢子叶球序,并在大、小孢子叶球的基部有多数不育的苞片;胚珠顶生,珠心和珠被完全分离(图13-43)。从上述特征可以看出,苛得荻植物在胚珠结构、叶的形态与结构等方面与种子蕨相似,而茎的构造和孢子叶的形态等又类似现有的裸子植物。
种子蕨和苛得荻之间并不存在系统发育上的祖裔关系,它们都是原裸子植物的后裔。根据现有的裸子植物化石资料,现存的裸子植物都是由原裸子植物沿两个方向演化而来,一是由古蕨经过复杂的分枝和次生组织的发育,在石炭纪形成苛得荻,再进一步发展成为银杏类、松柏类和红豆杉类,现存的裸子植物大多属于此类;另一支则由无脉蕨经过侧枝的简化,形成种子蕨,再进一步发展成为拟苏铁类(Cycadeoides)(图13-44)和苏铁类(Cycads),其中拟苏铁类在白垩纪后期绝灭。至于买麻藤纲植物的起源和系统地位,至今尚存有争议,根据它们形体的结构和明显的分节,被认为与木贼类植物有一定的亲缘关系;但从它们孢子叶球的结构来看,其祖先曾具有两性的孢子叶球,而具有两性孢子叶球的植物,只有起源于种子蕨的拟苏铁类,它们的孢子叶球二叉分枝和具有珠孔管等特点说明买麻藤纲植物很可能是强烈退化或特化了的拟苏铁植物的后裔,但买麻藤植物茎内维管组织具导管、精子无鞭毛、颈卵器趋于消失。以及类似花被结构(盖被)的形成和虫媒的传粉方式等,又是堪与被子植物相比拟的高级性状。
裸子植物的历史可远溯到3亿5千万年之前。化石研究表明,中泥盆纪的无脉蕨(Aneurophyton)(图13-41A)是原始裸子植物的一个代表,它是一种高大的乔木,茎顶端有一个由许多分枝组成的树冠,其末级“细枝”的形状就像分叉的叶片,但其中无叶脉;孢子囊小而呈卵形,生于末级“细枝”之上;茎干内部具次生木质组织,这种组织由带具缘纹孔的管胞组成;它没有发达的主根,只有许多细弱的侧根。古蕨(Archaeopteris)(图13-41B)是晚泥盆纪特有的一群较为进化的原始裸子植物的代表,它是高达18m以上的塔形乔木,茎的最大直径为1.5m,有形成层及次生组织,木质部成分有具缘纹孔的管胞,茎干的顶端有一个由枝叶组成的树冠;叶是扁平而宽大的羽状复叶;根系较无脉蕨发达;孢子囊单个或成束地着生在不具叶片的小羽片上,孢子囊内曾发现大、小两种孢子。尽管古蕨仍是以孢子进行繁殖的,但它的外部形态、内部结构和生殖器官的特征更接近裸子植物,因而推测它可能是由原始蕨向裸子植物演化的一个早期阶段或过渡类型,所以人们称古蕨为原裸子植物(Progymnospermae)或半裸子植物。到了晚泥盆纪、早石炭纪时,由原裸子植物演化出更高级的类型——种子蕨(Pteridospermae)、苏铁蕨(cycadofilices)和苛得荻(Cordaitinae)等。
种子蕨是一种最原始的种子植物,最早出现于早石炭纪的地层中,在晚石炭纪和二叠纪得到了极大发展,是当时陆生植被中的优势类群。种子蕨植物体不很高大,主茎很少分枝,叶为多回羽状复叶;种子小型,并有一杯状包被,其上生有腺体,种子中央为一颇大的雌配子体组织和颈卵器,珠心(大孢子囊)的顶端有一突出的喙,喙外又有一垣围之,两者之间为花粉室,其中有时可看见花粉粒,珠心之外有一厚的珠被(图13-42)。从上述特征可以看出,种子蕨是介于真蕨类植物和种子植物之间的一个过渡类型,但种子蕨并不是从真蕨起源的,而是从起源于裸蕨的原裸子植物演化而来。
注:本文部分资料来源于扬州大学生物系教材