一直以为cDNA是双链,老师问我生化是怎么学的?哎,底子薄终究是站不住脚的。现将这方面的知识温习一下。
真核生物的mRNA的3'末端具有一段大约150-200bp的多聚腺苷酸尾巴(原核生物没有3’端A尾巴,或者很短),该结构能够保护mRNA免受核酸外切酶的攻击,同时能够终结转录以及将mRNA从细胞核输出并进行翻译都起着十分重要的作用。
cDNA的概念简单来说是以RNA为模板,在逆转录酶的作用下,合成与模板RNA互补的单链DNA。模板RNA可以是总RNA,mRNA,以及体外转录的RNA产物,但无论是哪种,要确保没有RNA酶和DNA的污染。对于原核生物,因为没有多聚A尾,因此只能用随机引物(主要是色氨酸tRNA,6-10个核苷酸),在RNA逆转录酶(依赖RNA的DNA聚合酶)的催化作用下从5'→3'端合成与模板RNA互补的cDNA单链,形成RNA-DNA杂交双链。对于真核生物,除了可以用随机引物,还可以用oligoT(多聚胸腺嘧啶寡核苷酸链)作为引物与模板的3’端结合,从而形成杂交双链。
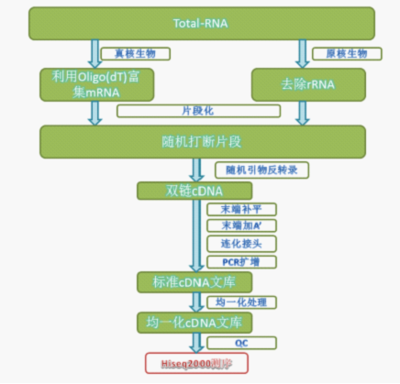
cDNA的第二条链是如何形成的呢?一种情况是利用 cDNA 第一链的 3'末端常常出现发夹环的特征,这种发夹结构是反转录酶在第一链末端“返折”并且进行复制第一链的结果,它为合成 cDNA第二链提供了有用的引物。用这种方法合成的双链 cDNA 在一端有一个发夹环,可以用单链特异的 S1 核酸酶切去。但是 S1核酸酶的处理,常常会“修剪 ” 过多的 cDNA 顺序,使 cDNA 丢失了 mRNA 5' 端的部分顺序。 另一种方法是用大肠杆菌的RNase H 进行修饰。 RNase H 能识别 RNA-DNA杂交分子并把其中的 RNA 切割成短的片段,这些 RNA 短片段仍与cDNA 第一链结合,可被新合成的 DNA 所取代。新合成的 DNA 存在切口,用 DNA 连接酶把这些切口连接在一起形成一条完整的DNA 链。 RNase H 法优于 S1 核酸酶法,它能获得包括 mRNA 5' 端全部或绝大部分的更长顺序 cDNA分子。
在以cDNA为模板的时候,为什么要设计正反两条引物呢?第一回合pcr以cDNA单链为模板,退火时正向引物与模板结合,从而合成与模板互补的第二条cDNA链。第二轮pcr开始时,两条引物均发挥作用,产物成指数增长。正向引物可与oligoT配对形成双引物,但反向引物和oligoT就不可以,原因是反向引物和oligoT方向相同,可以看做是同一个引物。