罂粟用于减轻疼痛已有近千年的历史。1803年由罂粟生物碱分离物质出的晶体,被证实是天然阿片的镇痛活性成份,称为吗啡。吗啡的立体化学结构是其与机休特异部位相互作用产生镇痛所必须。通过吗啡、酮唑辛和SKF-10047等一组激动药所产不同药理活性,确定了三种阿片类药物综合征,分别命名为m,k 和 s原型,由此导致了m, k 和s三种阿片受体的发现。后来发现与SKF-10047相关的s型综合征不能被普通阿片拮抗剂纳洛酮(naloxone)所阻断,因此s型受体不再被认为是阿片受体家族的成员。δ型受体是由kosterlitz小组在研究内源性阿片肽和内啡肽的效应时发现的。经过近30年的实验室研究,对m、k和δ型受体的认识已较清楚,其基因编码已被克隆,这3种受体称为“经典型阿片受体”。最近 cDNA编码一种称之为“孤立阿片”受体,经签定与经典阿片受体有高度同源性,它的结构基团是阿片受体,因此称其为阿片样受体(opioidreceptor-like,ORL1)。有药理学迹象表明每种阿片受体存在亚型,以及其他新型、较少了解的阿片受体e、l、i和z。本文着重介绍阿片受体研究进展。
一.经典阿片受体
三种经典m、k和δ阿片受体被确认后,发现在脑内分布广泛但不均匀。这些受体分布在痛觉传导区以及与情绪和行为有关的区域,集中分布在导水管周围灰质、内侧丘脑、杏仁核和脊髓胶质区。这些复杂的受体可以被不同的激动剂激活,产生不同的生物效应。例如主要分布于脑干的μ受体被吗啡激活后,可产生镇痛和呼吸抑制等作用,而主要分布于大脑皮质的κ受体只产生镇痛作用而不抑制呼吸。然而不同阿片受体在中枢神经系统的分布,以及对不同阿片配体结合能力存在差异。阿片受体的内源性配体为脑啡肽、内啡肽和强啡肽,它们分别由不同的基因编码。这些五肽对阿片受体的亲和力不同,但三者均可与一种以上的阿片受体结合。其中脑啡肽对δ型受体有较强的选择性,被认为是其内源性配体。强啡肽对k型受体选择性较强,是其内源性配体。m型受体的内源性配体直到1997年才被发现,称为内啡肽或内源性吗啡(endomorphine)。内源性吗啡在中枢神经系统与μ-阿片受体呈镜像分布,对μ受体的结合力比对δ和κ受体的结合力高100倍。
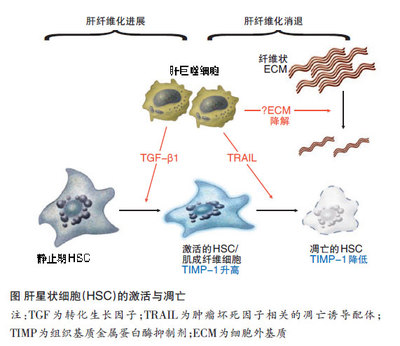
最近人们克隆出三种典型的阿片类受体μ、δ和κ,并确定了其核苷酸序列。阿片受体属于G蛋白耦联受体大类,该类受体具有相同的基本结构:一个细胞外氨基端区域,七个跨膜域以及一个细胞内羧基端尾区(如图-1所示)。
图1阿片受体基本结构示意图
克隆得到的阿片类受体有高度内源性,65%的氨基酸序列是相似的。最大的不同在于细胞外环、氨基端和羧基端区域。阿片受体的配体是二价的,一部分调节信号的传导,另一部分决定受体的选择性。它们分别称为信号区和结合区。信号的传导与跨膜域有关,而细胞外环的作用与配体从结合部位分离有关。μ受体和δ受体的第一个细胞外环只有7个氨基酸不同,两者最主要的区别在于108位点的一个氨基酸差异。δ受体中该位点的缬氨酸被门冬酰胺替换后,即可与μ受体配体DAMGO以高亲和力结合。受体的结合隐穴由多个跨膜域和细胞外环的多氨基酸立体结构形成。拮抗剂与受体结合后,不产生激活受体所需的构像改变。但单个氨基酸的改变(如TM4区亮氨酸取代丝氨酸)可使桔抗剂与受体作用后产生激动剂的作用。
二.经典阿片受体亚型
1.m-受体亚型Pasternak等采用放射性配体研究,以[3H]-标记-m、d和k受体配体,显示双向结合的特性。根据其研究结果将m受体分为m1和m2两个亚型,每种放射性配体m1受体具有相同的高亲和力,而不同的放射性配体则分别对m、d和k受体有不同的亲和力。纳洛刹腙(Naloxazone)和纳洛肼(naloxonazine)可取消每种放射性配体与m1受体的结合。此外,在活体研究中发现纳洛刹腙可选择性地阻断吗啡诱发的抗伤害作用,而不能阻断吗啡诱发的呼吸抑制和吗啡依赖作用。
有研究提示可能存在一种不同于m1和m2的新型m受体。与这种新型m受体结合的激动剂是吗啡结构环中的位置被置换的吗啡类似物,如吗啡-6b-葡萄糖醛酸苷、海洛因和6-乙酰吗啡,而吗啡本身并不与该受体相互作用。在鼠的抗伤害试验中,吗啡与吗啡-6b-葡萄糖醛酸苷、海洛因和6-乙酰吗啡均有较强的抗伤害性作用,而与吗啡之间并没有交叉耐受性。这些激动剂并不是通过d-或k-受体产生抗伤害性作用,因此它们的抗伤害性作用不能被选择性d或k-受体阻断剂所阻断。有报道3-甲氧纳曲酮可拮抗吗啡-6b-葡萄糖醛酸苷、海洛因和6-乙酰吗啡产生的抗伤害作用,而不影响产生的抗伤害作用。最近有研究显示,内源性吗啡-1基因外显子1被破坏,吗啡-6b-葡萄糖醛酸苷和海洛因仍可能产抗伤害作用,而吗啡则不能产生抗伤害作用。同一研究者在另一内源性吗啡-1基因外显子2被破坏的鼠,所有三种激动剂均不能产生抗伤害作用。因此得出结论,吗啡-6b-葡萄糖醛酸苷和海洛因在外显子1MOR-1变异鼠的抗伤害性作用是通过MOR-1基因转录替代物受体介导,这种受体不同于m1和m2的新型m-受体。
2.d-受体亚型 目前为止,被克隆d-受体基因只有DOR-1基因。然而,根据在体和离体的药理学实验,d-受体的两个亚型(d1/d2和dcx/dncx)有相互重叠。根据在体的药理学实验时,d受体配体的不同将d-受体分为两个亚型,即d1和d2 受体。在啮齿类在体实验中,DPDPE的脊髓上的抗伤害性作用可被7-苯亚甲基纳曲酮(BNTX)或DALCE 选择性拮抗。然而,δ啡肽Ⅱ和脑啡肽的抗伤害作用可被naltriben或 naltrindole所逆转。给鼠反复注射DPDPE或δ啡肽Ⅱ产生的对抗伤害效应的耐受,而对不同配体所引起的耐受之间没有交叉。在体的实验显示d1和d2-受体诱发的抗伤害效应可被不同类型的钾离子通道阻滞剂所拮抗22。实际上,DPDPE可能不仅是一个选择性的d1受体激动剂,也可能是d2-受体的部分激动剂。
d-受体还存在另外亚型,即dcx和dncx受体。dcx受体是一种复合有m-受体(可能还有k-受体)的d受体亚型。而dncx受体并不与另一种阿片受体相联系。亚抗伤害剂量的dcx受体激动剂(如低剂量DPDPE),可以增强m-受体介导的镇痛作用。另一方面,高剂量DPDPE可以作为dncx-受体激动剂,本身可诱发镇痛作用,这一作用可被DALCE逆转。后来通过放射性配体研究证实dncx受体又有d(ncx-1) 和d(ncx-2).亚型之分。最近研究提示d(ncx-1)受体和dcx-受体实际分别与以前的分类d1-受体和d2-受体同义。
3.k-受体亚型采用[3H]-乙基氯代环唑星(EKC)研究豚鼠脊髓k-受体,首次确认了k1和k2-受体亚型的存在,并逐渐被接受。进一步研究显示还有k3-受体亚型的存在,k3-受体亚型介导丙烯吗啡的抗伤害效应。选择性k受体激动剂U-50,488和其同源物(U-69,593,PD117302, CI 977, ICI197067)在研究中的应用,使得k受体的分型更加精确。k1和k2受体亚型又进一步分为k1a、k1b和k2a、k2b。由于缺乏特异性的k-受体亚型的拮抗剂,还没有确切功能性药理学的证据k-受体各亚型的存在。然而有报道在鼠用U-50,488类似物异硫氰酸盐预处理后,能长时间阻断U-69,593的抗伤害效应,而不影响布马佐辛的抗伤害作用。用非选择性拮抗剂夸达佐辛预处理,则可选择性地阻断马佐辛的抗伤害作用。这些研究结果都有力地支持k-受体分为k1和k2两个亚型。
三.阿片样受体(ORL1)
阿片受体被克隆后不久,学者们又分离出一种具有G-蛋白-耦联受体典型结构的蛋白,这种蛋白与经典阿片受体的结构具有同源性,被接受为阿片受体家族的成员,称为阿片样受体(ORL1)。但ORL1与经典阿片受体没有相应的药理学同源性,甚至对m-,k- 和d-受体有高度均一亲和力的非选择性配体,对ORL1受体也只有很低的亲和力。由于这个原因,开始没有发现ORL1的内源性配体,故称其为“孤立阿片受体”。“孤立阿片受体”并没有孤立很长时间,不久确认新发现的70肽菌素为其内源性配体,并被命名为伤害感受素(nociceptin),其序列的起始端和结束端氨基酸分别为苯丙氨酸(F)和谷氨酰胺(Q),又称之为FQ孤啡肽。FQ孤啡肽引起的细胞反应与经型阿片受体激活反应相似。动物的伤害感受素则产生一系列与其他阿片类不同的生物效应。鞘内注射伤害感受素可产生镇痛作用,但脑室内给药则引起痛觉过敏并拮抗阿片类镇痛作用。伤害感受素刺激摄食、诱发焦虑,并参与记忆和中枢信息加工。
虽然有些研究结果提示有ORL1亚型的存在,但现在就作出此结论还未时尚早。最可靠的受体药理学定义需根据受体拮抗剂亲和力的差异,而对于ORL1受体缺乏有用的拮抗剂。尽管有伤害感受素N-末端十三肽的合成类似物可以作为选择性拮抗剂,当增加用量时又倾向于产生激动作用。同时也没有理由说这个肽对外周的ORL1受体具有抗作用。
四.其他阿片受体
除了m-, d-, k-和ORL1-受体外,推测还有几种其他阿片受体,如e-, l-, i-和z受体。e-受体,对b-内啡肽具有特异性。兔回肠有i-受体,对脑啡肽有高度亲和力但又不同与d-受体。在新鲜鼠膜碎片有一种很不稳l结合位点,对4,5-环氧左马南有高度亲和力。对e-,l-, i-和z-受体特性了解的很少。只有得到进一步的实验证据,尤其是它们特异性cDNAs得到分离,这些阿片受体才能得到更广泛的认可。
五.外周阿片受体
阿片受体长期以来被认分布于中枢神经系统。近十多年研究表明在中枢神经系统外也存在阿片受体,这些受体统称为外周阿片受体。解剖学、分子学和电生理学研究显示三种经典阿片受体(μ,δ,κ)在感觉神经元、背根神经节细胞和初级传入神经元末均有分布。在Meissner小体内的传入纤维和无髓鞘感觉神经的末梢可找到阿片受体,但在交感节后神经元上无分布。外周阿片受体介导的镇痛作用对动物和人的炎性疼痛尤其显著。外周感觉神经的阿片受体受炎症的精密调控。位于炎性组织范围内的免疫细胞表达这些受体的内源性配体称为阿片肽,局部刺激以及释放剂能使阿片肽释放引起局部镇痛,免疫系统受抑制可阻断这种作用。因此认为内源性阿片肽由免疫细胞分泌,它们与感觉神经上的阿片受体结合,通过抑制这些神经的兴奋性和(或)炎症前神经肽的释放而产生镇痛作用。
在受伤组织局部给予小剂量经典阿片受体激动剂,并不能激动中枢神经系统的阿片受体,但可通过外周阿片受体介导而产生镇痛效果。当阿片受体激动剂结合一个具有高度极性亲水性的取代基后,可限制其进入中枢神经系统。洛哌丁胺是一种止泻药,对克隆的人μ阿片受体有高度亲和性和选择性,是μ受体激动剂。由于洛哌丁胺对于脂膜有高度亲和性,并且有减低表面张力的能力,在膜上积聚而不吸收至全身。因此其可作为外周μ受体激动剂,对于热伤、表皮损伤处、甲醛、弗洛依德左剂引起的炎症和膝关节炎局部应用后起作用。静脉使用或口服后,可以分别在肝、肾、肺或胃、肠中积聚。阿西马多林是一种有效的选择性κ激动剂,几乎不通过血脑屏障。
阿片类药物全身应用常伴有副作用,如瘙痒、尿潴留、恶心呕吐、胃排空延迟以及便秘等,当镇痛作用消失后,其副作用有可能仍然存在。阿片类药物的副作用常是由于其作用于外周阿片受体所引起,如使用外周阿片受体拮抗剂,可特异性的减弱阿片类药物的外周副作用,而保持中枢镇痛作用保持不变。口服和皮下给予外周阿片受体拮抗剂甲基纳曲酮,可以减轻阿片相关的瘙痒及烦躁,迅速逆转吗啡引起的胃排空延迟。
六.阿片受体激动的效应
阿片受体属G蛋白偶联受体(GPCR),激动后一般产生抑制作用。当激动剂与阿片受体结合后激活Gi蛋白,使G蛋白的βγ亚基与α亚基解离。βγ亚基与α亚基分别介导了【】胞内多条信号通路的激活,如腺苷酸环化酶活性的抑制、G蛋白偶联受体激酶(GRK)、PKC和MAPK的激活等。从而关闭N型电压控制型钙通道,开放钙依赖性内控型钾通道。由此导致超极化和神经元兴奋性下降。纳摩尔级的阿片类药物并不产生抑制作用,而是通过激活兴奋性Gs蛋白而产生兴奋作用,正因为这种原因,合用极低剂量的拮抗剂可显著增强阿片类激动剂的镇痛作用。使用极低剂量长效拮抗剂纳洛酮或纳美酮芬(nalmefene)可显著降低术后患者的吗啡用量及呕吐、瘙痒等副作用的发生率。
经典的m-, k- 和 d-受体阿片受均有明确内源性配体,激动后产生的效应如表-1所示。
表1 阿片受体配基及激动效应
受体 | 激动剂 | 拮抗剂 | 激动效应 |
μ | 内啡肽 | 纳洛酮 | 脊髓上水平镇痛 |
吗啡 | 纳曲酮 | 呼吸抑制 | |
芬太尼 | 瞳孔缩小 | ||
舒芬太尼 | 抑制胃肠蠕动 | ||
哌替啶 | 恶心、呕吐 | ||
δ | 脑啡肽 | 纳洛酮 | 脊髓水平镇痛 |
DPDPE | IC1154,126 | 抗镇痛 | |
DADLE | |||
k | 强啡肽 | 纳洛酮 | 脊髓水平镇痛 |
U 50,488 | MR2266 | 瞳孔缩小(弱) | |
呼吸抑制(弱) | |||
焦虑 |
由于内源性配体其在体内很快被降解或不能通过血脑屏障,因此不能用于临床治疗目的。不同阿片类镇痛药与各种阿片受体有着不同的亲和力,产生不同的药理效应而广泛用于临床。根据与阿片受体的亲和力的不同,可将阿片类药物分为激动剂、部分激动剂、激动—拮抗混合剂、拮抗剂。阿片类药物的镇痛作用主要是激动m1受体,m2受体激动主要与不良反应相关。现有的阿片类药物对m1受体的选择性无明显差别。因此,现有的强阿片类药物作用机制相似,不良反应也相似,而吗啡是其中最经典强阿片类药物。未来药物研究方向是寻找选择性作用于m1受体的药物。传统认为δ受体激动也主要产生镇痛作用,但与μ受体激动后产生镇痛作用不同的是,μ受体激动后患者感觉舒畅和愉悦,而δ受体激动后患者会出现躁动难受。最近上海生命科学研究院神经科学研究所通过大量研究揭开了δ受体的两个两个谜:一是它潜藏在细胞内部。而此前一直认为它同μ阿片受体一样,也是分布于神经细胞的表面。二是发现δ受体是柄"双刃剑",既镇痛,又致痛,因为运载受体的囊泡内含有致痛物质。当它受到疼痛或致痛药物的刺激后,由细胞内部到细胞表面镇痛。然而它在镇痛的同时,内部的致痛物质也随之而出。这一发现,给临床麻醉和镇痛治疗提供了新的理论基础和用药新思路:在激活μ受体进行镇痛的同时,可用药物阻断δ受体,这样既可减少阿片类镇痛药用量,又可提高镇痛效果,同时又减少阿片类镇痛药的副作用。
总之,阿片受体自发现以来,一直是生命科学的研究热点之一,阿片类药物的镇痛、耐受及成瘾机制的阐明以及新的更理想阿片类药物的研制,这些都离不开对阿片受体及相关蛋白的不断深入研究。
(上海第二医科大学附属瑞金医院麻醉科 彭章龙)