buqinhexing
不亲和性
incompatibility
在过程中由于生物个体的细胞或组织水平上的不协调而使受精或接合不能正常进行,或是受精后不能产生后代的现象。这是双亲个体间基因型的不协调或是染色体组非同源性在生殖生理表型上的一种反映。不亲和性是稳定物种的重要机理之一,也有利于维持群体中个体的杂合性,因而对于物种的生存和进化有一定的意义。
1877年英国生物学家首先描述了生殖不亲和现象。他发现樱草的花有所谓针式和线式两种。针式花具有长的雌蕊和短的雄蕊,线式花具有短的雌蕊和长的雄蕊。只有针式花的花粉落在线式花的柱头上或是线式花的花粉落在针式花的柱头上才能顺利地发芽受精。因此樱草种内的个体按花器形态可分属于两个交配型,交配型之间是交配亲和的,交配型之内不仅自交不亲和,而且异交也不亲和。后来发现类似的不亲和现象在同型花植物中也广泛存在。异形花蕊型不亲和性植物有荞麦、干屈菜、亚麻、连翘等;同形花蕊型不亲和性植物有甘蓝、白菜、矮牵牛、月见草、漆姑草等。在低等的雌雄同体动物中也常有特殊的自交不孕现象。达尔文认为这些特性都是的结果。
类型 种间不亲和性 异种、异属生物个体之间的生殖不亲和现象常由染色体组的不同所造成。一般地说,越是远缘则不亲和性越高,直到完全不能受精。但也有分类系统上关系较远的个体可以,而比较近缘的反而不亲和的例子。例如马和驴可产生骡驹,而山羊和绵羊却很难杂交。
种内不亲和性 种内个体间的不亲和现象往往是受不亲和基因所制约而并不是染色体组不同的结果。它可分为:①自交不亲和性,包括同株异花间授粉不亲和,同花雌雄蕊之间授粉不亲和以及同一无性繁殖系内个体间授粉不亲和等情况;②异交不亲和性,自交不亲和的植株与不属于同一无性繁殖系的其他植株交配时仍表现为不亲和的现象。
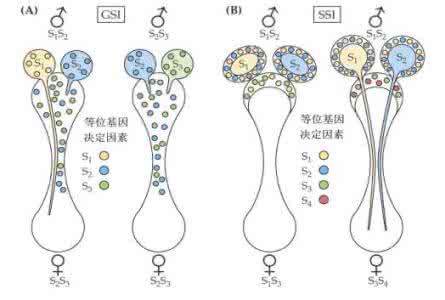
不亲和基因 按所处的部位可分为两类:一类是自交不亲和的染色体基因。1915年曾由美国学者E.M.伊斯特在烟草属中发现 3个复等位的不亲和染色体基因S、S、S。授粉时如花粉中的不亲和基因与花柱基因型中所包含的某个不亲和基因相同时便不能受精。在S、S、S之间没有显隐性关系,因此SS(♀)×SS(♂)的组合中只有S花粉能顺利地受精, 而反交SS(♀)×SS(♂)则完全不亲和(表1[不亲和基因在不同的交配组合中的相互关系]) 。另一类是不亲和的染色体外基因。许多高等植物的某些雄性不育现象是染色体外基因和染色体基因不亲和的结果。在玉米等植物中, 染色体外基因是 S而染色体基因是FF或Ff的雄性植株能产生正常的花粉,是雄性可育的,说明染色体外基因S和染色体基因F是亲和的。染色体外基因是 S而染色体基因是ff的个体产生败育的花粉,是雄性不育的,说明染色体外基因S和染色体基因 f不亲和。雄性不育的个体本身不能保持后代。因此必须借助于另一类染色体基因是ff而染色体外基因是N的保持系才能保种繁殖。
不同的不亲和体系中的不亲和基因的数目常不相同。不亲和性由一对基因所决定的体系称为二极型,例如衣藻、链孢霉的交配型只有正(+)、负(-)之分。不亲和性由两对基因所决定的体系称为四极型,存在于菌类和某些禾本科植物中。不亲和基因座位愈多,个体间的亲和性选择就愈严格。一对不亲和基因的生物的后代亲和的组合占总杂交组合的1/2,两对不亲和基因是1/4(表2[两对基因支配的不亲和性]),对不亲和基因是。一个不亲和性座位上可以有两个以上的等位基因,例如烟草的不亲和性座位上至少有15个复等位基因,月见草有45个,而某些真菌可达100个以上。
从种子植物不亲和基因作用的自主性来看,不亲和性又可分为①配子型不亲和性,即自主性的不亲和性不亲和性完全取决于花粉的不亲和基因与花柱的不亲和基因是否相同(表1[不亲和基因在不同的交配组合中的相互关系])②孢子型不亲和性,即非自主性的不亲和性,花粉的表型受花粉赖以生长发育的孢子体的基因型所制约。例如在某些菊科和十字花科植物中, SS(♀)×SS(♂)组合中的S是显性的,所以S花粉在SS的花柱上和S花粉一样会发芽、受精。孢子型不亲和植物多数是异形花蕊型。樱草的孢子型不亲和性由一对不亲和基因S和s所控制,另一对等位基因G和 g决定雌蕊长短(G是短雌蕊,g是长雌蕊),等位基因A和a决定雄蕊长短(A是长雄蕊,a是短雄蕊),这三个基因紧密连锁而且极少发生交换。针式花型的基因型是纯合型 gsa/gsa;线式花型的基因型是杂合型GSA/gsa,线式孢子体产生的gsa花粉与针式的gsa花柱是亲和的,而与线式的GSA花柱则是不亲和的。这样由不亲和基因所导致的杂交结果使群体后代中针式花型的纯合基因型与线式花型的杂合基因型永远能维持1与1之比。
生
理机制 种子植物的生殖不亲和反应或是由于花粉在柱头上不能发芽,或是由于在花柱中的伸长延迟所造成。按照花粉在雌蕊中受抑制的部位,不亲和反应可分为三类:柱头类、花柱类和子房类。孢子型不亲和反应多属柱头类,不亲和反应发生在柱头的乳头细胞上;配子型的不亲和反应多数发生在花柱中,属花柱类,表现为花粉管伸长延缓,直到最后停止;不亲和反应如果发生在子房中则称为子房类。
实践意义 育种 种间不亲和性带来远缘杂交的困难,自交不亲和性带来保持纯系的困难,因此如何克服不亲和性是育种工作中的一项研究课题。在十字花科中一般采用蕾期授粉法(即小花花冠未展放之前进行人工授粉)以及对位于二次分枝末端部位的小花进行授粉的方法。用环己胺处理芸苔属花粉也有显著效果。在胡萝卜中可采用切去柱头或在花柱基部打孔再在孔穴内置入花粉的授粉法来克服自交不亲和性。这些都是针对不亲和物质可能存在的部位及其转移方向而采取的措施。自交不亲和性极强的月见草雌蕊用 50℃高温处理5分钟就可以克服不亲和性。此外辅助混合授粉法,即将不亲和花粉辅以失活的亲和花粉或其匀浆一起授粉也可收到一定的效果。
辐射处理早期的花粉母细胞能使某些自交不亲和植物发生突变而成为自交亲和的,这在欧洲甜樱桃和一种月见草中都有报道。在某些场合(如樱草)多倍化也有降低自交不亲和的效应。例如四倍体的基因型是SSSS,则花粉的基因型是SS、SS、SS,其中杂合基因型的花粉在某些植物中有某种程度的自交结实或异交结实能力。
属于染色体不亲和的远缘种之间可借助中间类型媒介法、混合授粉法、染色体加倍法、试管受精法、杂种胚人工培养法等求得某种程度的杂交可能性。遗传工程则是近年发展起来的一种克服种间不亲和性的有效方法。
进化研究 植物在自然选择过程中形成的不亲和性,特别是自交不亲和性与动植物个体的性别分化相似,都有保持基因杂合性的效果。不亲和性又是保证物种平衡和稳定的重要因素之一,它有利于维持个体基因成分的杂合性,能提高物种本身的生存竞争能力。在微生物中,异宗结合往往与感染力有关。例如含有不同交配型(+和-)的锈病菌和黑锈病菌的异核体可使寄主致病,然而单核体菌丝或偶尔形成的同型双核体 (++,或──)全部都是非寄生的。这里寄生性完全取决于不亲和基因的杂合性;不亲和基因同样提高了锈病菌的生存竞争能力。
葛扣麟
以上就是网友分享的关于"不亲和性"的相关资料,希望对您有所帮助,感谢您对爱华网的支持!