核糖核酸(英语:Ribonucleic acid,缩写:RNA)存在于生物细胞以及部分病毒、类病毒中的遗传信息载体。由至少几十个核糖核苷酸通过磷酸二酯键连接而成的一类核酸,因含核糖而得名,简称RNA。每个RNA分子都由核苷酸单元长链组成,每个核苷酸单元含有一个含氮碱基、一个核糖和一个磷酸基。RNA是具有细胞结构的生物的遗传讯息中间载体,并参与蛋白质合成;还参与基因表达调控。对一部分病毒而言,RNA是其唯一的遗传物质。RNA存在于大多数已知的植物病毒和部分动物病毒以及一些噬菌体中。
核糖核酸_核糖核酸 -分类
核糖核酸
RNA是以DNA的一条链为模板,以碱基互补配对原则,转录而形成的一条单链,主要功能是实现遗传信息在蛋白质上的表达,是遗传信息传递过程中的桥梁。tRNA的功能是携带符合要求的氨基酸,以mRNA为模板,合成蛋白质。
RNA由核糖核苷酸经磷酯键缩合而成长链状分子。一个核糖核苷酸分子由磷酸,核糖和碱基构成。RNA的碱基主要有4种,即A腺嘌呤
,G鸟嘌呤
,C胞嘧啶
,U尿嘧啶
。其中,U尿嘧啶取代了DNA中的T胸腺嘧啶而成为RNA的特征碱基。
mRNA
mRNA的功能就是把DNA上的遗传信息精确无误地转录下来,然后再由mRNA的碱基顺序决定蛋白质的氨基酸顺序,完成基因表过程中的遗传信息传递过程。在真核生物中,转录形成的前体RNA中含有大量非编码序列,大约只有25%序列经加工成为mRNA,最后翻译为蛋白质。因为这种未经加工的前体mRNA(pre-mRNA)在分子大小上差别很大,所以通常称为不均一核RNA(heterogeneousnuclearRNA,hnRNA)。
tRNA
如果说mRNA是合成蛋白质的蓝图,则核糖体是合成蛋白质的工厂。但是,合成蛋白质的原材料――20种氨基酸与mRNA的碱基之间缺乏特殊的亲和力。因此,必须用一种特殊的RNA――转移RNA(transferRNA,tRNA)把氨基酸搬运到核糖体上,tRNA能根据mRNA的遗传密码依次准确地将它携带的氨基酸连结起来形成多肽链。每种氨基酸可与1-4种tRNA相结合,已知的tRNA的种类在40种以上。
tRNA
tRNA是分子最小的RNA,其分子量平均约为27000(25000-30000),由70到90个核苷酸组成。而且具有稀有碱基的特点,稀有碱基除假尿嘧啶核苷与次黄嘌呤核苷外,主要是甲基化了的嘌呤和嘧啶
。这类稀有碱基一般是在转录后,经过特殊的修饰而成的。
1969年以来,研究了来自各种不同生物,:如酵母、大肠杆菌、小麦、鼠等十几种tRNA的结构,证明它们的碱基序列都能折叠成三叶草形二级结构(图3-23),而且都具有如下的共性:
①5’末端具有G(大部分)或C。
②3’末端都以ACC的顺序终结。
③有一个富有鸟嘌呤的环。
④有一个反密码子环,在这一环的顶端有三个暴露的碱基,称为反密码子(anticodon).反密码子可以与mRNA链上互补的密码子配对。
⑤有一个胸腺嘧啶环。
rRNA
核糖体RNA(ribosomalRNA,rRNA)是组成核糖体的主要成分。核糖体是合成蛋白质的工厂。在大肠杆菌中,rRNA量占细胞总RNA量的75%-85%,而tRNA占15%,mRNA仅占3-5%。
rRNA
rRNA一般与核糖体蛋白质结合在一起,形成核糖体(ribosome),如果把rRNA从核糖体上除
掉,核糖体的结构就会发生塌陷。原核生物的核糖体所含的rRNA有5S、16S及23S三种。
S为沉降系数(sedimentationcoefficient),当用超速离心测定一个粒子的沉淀速度时,此速度与粒子的大小直径成比例。5S含有120个核苷酸,16S含有1540个核苷酸,而23S含有2900个核苷酸。而真核生物有4种rRNA,它们分子大小分别是5S、5.8S、18S和28S,分别具有大约120、160、1900和4700个核苷酸。rRNA是单链,它包含不等量的A与U、G与C,但是有广泛的双链区域。在双链区,碱基因氢键相连,表现为发夹式螺旋。
rRNA在蛋白质合成中的功能尚未完全明了。但16S的rRNA3’端有一段核苷酸序列与mRNA的前导序列是互补的,这可能有助于mRNA与核糖体的结合。
miRNA
miRNA
MicroRNAs(miRNAs)是在真核生物中发现的一类内源性的具有
调控功能的非编码RNA,其大小长约20~25个核苷酸。成熟的miRNAs是由较长的初级转录物经过一系列核酸酶的剪切加工而产生的,随后组装进RNA诱导的沉默复合体,通过碱基互补配对的方式识别靶mRNA,并根据互补程度的不同指导沉默复合体降解靶mRNA或者阻遏靶mRNA的翻译。最近的研究表明miRNA参与各种各样的调节途径,包括发育、病毒防御、造血过程、器官形成、细胞增殖和凋亡、脂肪代谢等等。
除了上述几种主要的RNA外还有一些其他RNA:
小分子RNA
(small RNA)
small RNA
存在于真核生物细胞核和细胞质中,它们的长度为100到300个碱基(酵母中最长的约1000个碱基)。多的每个细胞中可含有105 ~106 个这种RNA分子,少的则不可直接检测到, 它们由RNA聚合酶Ⅱ或RNA聚合酶Ⅲ所合成, 其中某些象mRNA一样可被加帽。
主要有两种类型的小分子RNA:一类是snRNA(small nuclear RNA),存在于细胞核中;另一类是scRNA(small cytoplasmic RNA),存在于细胞质中。
小分子RNA通常与蛋白质组成复合物, 在细胞的生命活动中起重要的作用, 。
①snRNA:
snRNA (smallnuclearRNA,小核RNA)。它是真核生物转录后加工过程中RNA剪接体(spilceosome)的主要成分。发现有五种snRNA,其长度在哺乳动物中约为100-215个核苷酸。snRNA一直存在于细胞核中,与40种左右的核内蛋白质共同组成RNA剪接体,在RNA转录后加工中起重要作用。某些snRNPs和剪接作用密切相关,它们分别与供体和受体剪接位点以及分支顺序相互补。
其中位于核仁内的snRNA称为核小体RNA(small uncleolar RNA),参与rRNA前体的加工及核糖体亚基的组装。
②scRNA:
scRNA(small cytoplasmic RNA,细胞质小RNA)主要位于细胞质内,种类较多,参与蛋白质的合成和运输。SRP颗粒就是一种由一个7SRNA和六种蛋白质组成的核糖核蛋白体颗粒,主要功能是识别信号肽, 并将核糖体引导到内质网。
端体酶RNA
端体酶RNA
端体酶RNA(telomeraseRNA),它与染色体末端的复制有关。
反义RNA
反义RNA(antisenseRNA),它参与基因表达的调控。
上述各种RNA分子均为转录的产物,mRNA最后翻译为蛋白质,而rRNA、tRNA及snRNA等并不携带翻译为蛋白质的信息,其终产物就是RNA。
核酶
另外还有一种特别的RNA(其分类与上述RNA分类无关)――核酶
核酶(ribozyme)一词用于描述具有催化活性的RNA, 即化学本质是核糖核酸(RNA), 却具有酶的催化功能。核酶的作用底物可以是不同的分子, 有些作用底物就是同一RNA分子中的某些部位。核酶的功能很多,有的能够切割RNA, 有的能够切割DNA, 有些还具有RNA 连接酶、磷酸酶等活性。与蛋白质酶相比,核酶的催化效率较低,是一种较为原始的催化酶。
大多数核酶通过催化转磷酸酯和磷酸二酯键水解反应参与RNA自身剪切、加工过程,也具有特异性,甚至具有Km值。
其发现是 科学家大肠杆菌RNaseP蛋白在切去部分后,在体外高浓度镁离子的情况下,留下的RNA部分(MIRNA)具有酶活性 。
非编码RNA
核糖核酸
【新型生命暗物质】非编码RNA(核糖核酸),被称为生命体中“暗物质”。日前,中国科学技术大学单革教授实验室发现一类新型环状非编码RNA,并揭示了此类非编码RNA的功能和功能机理。成果发表在国际知名杂志《自然・结构和分子生物学》上。非编码RNA是一大类不编码蛋白质,但在细胞中起着调控作用的RNA分子
。
正如宇宙间存在着许多既看不到也感觉不到的“暗物质”“暗能量”一样,在生命体这个“小宇宙”中,也存在这样的神秘“暗物质”―非编码RNA。
越来越多的证据表明,一系列重大疾病的发生发展与非编码RNA调控失衡相关。
环形RNA分子最近数年才引起研究人员注意,而此前的研究主要集中于线形RNA分子。单革教授实验室发现的新型环状非编码RNA,被命名为外显子-内含子环形RNA。在论文中,他们还对这类新型环状非编码RNA为何会成为环形而不是线形分子进行了研究,发现成环序列两端经常会有互补的重复序列存在
。
核糖核酸_核糖核酸 -细胞中的分布
蟾蜍血涂片
左图是用吡罗红甲基绿染色液染色的蟾蜍血涂片。
由于DNA和RNA在化学组成与分子结构上存在一定的差别,因而对不同的染料有着不同的反应。所以,可以根据这一反应差异来研究细胞中DNA与RNA的分布情况。
DNA和RNA两种核酸分子都是多聚体,但是它们的聚合程度有所不同。DNA聚合程度高,易于甲基绿结合
;RNA聚合程度低易于吡罗红结合
。所以当吡罗红与甲基绿混在一起作为染料时吡罗红与核仁、细胞质中的RNA选择性结合,从而显示红色;甲基绿与染色质中的DNA选择性结合,从而显示绿色。综上所述,RNA对吡罗红的亲和力大,被染成红色;DNA对甲基绿的亲和力大,被染成绿色。
核糖核酸_核糖核酸 -组成结构
与DNA不同,RNA一般为单链长分子,不形成双螺旋结构,
核糖核酸但是很多RNA也需要通过碱基配对原则形成一定的二级结构乃至三级结构来行使生物学功能。RNA的碱基配对规则基本和DNA相同,不过除了A-U、G-C配对外,G-U也可以配对。
在细胞中,根据结构功能的不同,RNA主要分三类,即tRNA(转运RNA),rRNA(核糖体RNA),mRNA(信使RNA)。mRNA是合成蛋白质的模板,内容按照细胞核中的DNA所转录;tRNA是mRNA上碱基序列(即遗传密码子)的识别者和氨基酸的转运者;rRNA是组成核糖体的组分,是蛋白质合成的工作场所。
在病毒方面,很多病毒只以RNA作为其唯一的遗传信息载体(有别于细胞生物普遍用双链DNA作载体)。
核糖核酸
1982年以来,研究表明,不少RNA,如I、II型内含子,RNaseP,HDV,核糖体大亚基RNA等等有催化生化反应过程的活性,即具有酶的活性,这类RNA被称为核酶(ribozyme)。
20世纪90年代以来,又发现了RNAi(RNAinterference,RNA干扰)等等现象,证明RNA在基因表达调控中起到重要作用。
在RNA病毒中,RNA是遗传物质,植物病毒总是含RNA。近些年在植物中陆续发现一些比病毒还小得多的浸染性致病因子,叫做类病毒。类病毒是不含蛋白质的闭环单链RNA分子,此外,真核细胞中还有两类RNA,即不均一核RNA(hnRNA)和小核RNA(snRNA)。hnRNA是mRNA的前体;snRNA参与hnRNA的剪接(一种加工过程)。自1965年酵母丙氨酸tRNA的碱基序列确定以后,RNA序列测定方法不断得到改进。除多种tRNA、5SrRNA、5.8SrRNA等较小的RNA外,尚有一些病毒RNA、mRNA及较大RNA的一级结构测定已完成,如噬菌体MS2RNA含3569个核苷酸。
核糖核酸_核糖核酸 -干扰机制
核糖核酸
1990年,曾有科学家给矮牵牛花插入一种催生红色素的基因,希望能够让花朵更鲜艳。但意想不到的事发生了:矮牵牛花完全褪色,花瓣变成了白色!科学界对此感到极度困
惑。
类似的谜团,直到美国科学家安德鲁・法尔和克雷格・梅洛发现核糖核酸
RNA(核糖核酸)干扰机制才得到科学的解释。两位科学家也正是因为1998年做出的这一发现而荣获2006年的诺贝尔生理学或医学奖。
上世纪八十年代,托马斯.R.切赫博士在研究RNA的成熟体结构中,发现了可以自我拼接的RNA催化作用(核糖核苷酸酶),并依此荣获1989年诺贝尔化学奖。经过多年的深度研究,切赫博士在DNA基因遗传过程中,发现了有趣的mRNA(信使RNA)和tRNA(转运RNA),从而揭开了遗传基因导致出生缺陷、大脑发育、营养吸收、细胞变异以及健康长寿等一系列人类生命密码的神秘面纱。
mRNA(信使RNA)人类的遗传信息主要贮存于DNA的碱基序列中,不过DNA并不直接决定蛋白质的合成。而在真核细胞中,DNA主要贮存于细胞核中的染色体上,而蛋白质的合成场所存在于细胞质中的核糖体上,因此需要有一种中介物质,才能把DNA 上控制蛋白质合成的遗传信息传递给核糖体。切赫博士把这种起着传递遗传信息作用的特殊RNA。称为信使RNA(messenger RNA,mRNA)。
简单的说,mRNA就是为了完成基因表达过程中的遗传信息传递。
令人遗憾的是,在遗传转录形成的过程中,仅有25%序列经加工成为mRNA,其余的均呈现非编码序列的前体mRNA形式,这些形势的mRNA在分子大小上差别很大,是导致出生缺陷、大脑发育、营养吸收、细胞变异以及健康长寿等一系列问题的基因遗传因素的关键所在。
切赫博士历经20年升华钻研,成果破译了mRNA编码序列信息奥秘,通过特殊的生物干预手段,优化mRNA的序列加工,筛查和剔除基因排列诱发基因和细胞突变的序列,不仅确保mRNA的序列加工的有效与增强,而且从根本上避免不良基因传递或传递序列问题引发细胞突变等一系列遗传问题的发生。
mRNA编码序列信息的成果破译,奠定了OMG配方盐技术的可行性基础。
法尔和梅洛的发现
科学家在矮牵牛花实验中所观察到的奇怪现象,其实是因为生物体内某种特定基因“沉默”了。导致基因“沉默”的机制就是RNA干扰机制。
此前,RNA分子只是被当作从DNA(脱氧核糖核酸)到蛋白质的“中间人”、将遗传信息从“蓝图”传到“工人”手中的“信使”。但法尔和梅洛的研究让人们认识到,RNA作用不可小视,它可以使特定基因开启、关闭、更活跃或更不活跃,从而影响生物的体型和发育等。
诺贝尔奖评审委员会在评价法尔和梅洛的研究成果时说:“他们的发现能解释许多令人困惑、相互矛盾的实验观察结果,并揭示了控制遗传信息流动的自然机制。这开启了一个新的研究领域。”
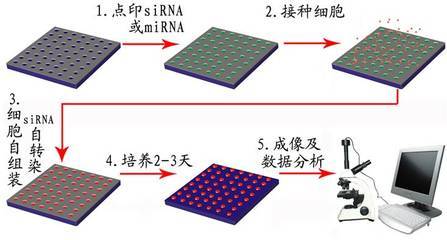
siRNA 的作用原理
核糖核酸
RNA干涉(RNAi)在实验室中是一种强大的实验工具,利用具有同源性的双链RNA(dsRNA)诱导序列特异的目标基因的沉寂,迅速阻断基因活性。siRNA在RNA沉寂通道中起中心作用,是对特定信使RNA(mRNA)进行降解的指导要素。siRNA是RNAi途径中的中间产物,是RNAi发挥效应所必需的因子。siRNA的形成主要由Dicer和Rde-1调控完成。由于RNA 病毒入侵、转座子转录、基因组中反向重复序列转录等原因,细胞中出现
了dsRNA,Rde-1(RNAi缺陷基因-1)编码的蛋白质识别外源dsRNA,当dsRNA达到一定量的时候,Rde-1引导dsRNA与Rde-1编码的Dicer(Dicer是一种RNaseIII 活性核酸内切酶,具有四个结构域:Argonaute家族的PAZ结构域,III型RNA酶活性区域,dsRNA结合区域以及DEAH/DEXHRNA解旋酶活性区)结合,形成酶-dsRNA复合体。在Dicer酶的作用下,细胞中的单链靶mRNA(与dsRNA具有同源序列)与dsRNA的正义链互换,原来dsRNA中的正义链被mRNA代替而从酶-dsRNA复合物中释放出来,然后,在ATP的参与下,细胞中存在的一种RNA诱导的沉默复合体RNA-induced silencing complex (RISC,由核酸内切酶、核酸外切酶、解旋酶等构成,作用是对靶mRNA进行识别和切割)利用结合在其上的核酸内切酶的活性来切割dsRNA上处于原来正义链位置的靶mRNA分子中与dsRNA反义链互补的区域,形成21-23nt的dsRNA小片段,这些小片段即为siRNA。RNAi干涉的关键步骤是组装RISC和合成介导特异性反应的siRNA蛋白。siRNA并入RISC中,然后与靶标基因编码区或UTR区完全配对,降解靶标基因,因此说siRNA只降解与其序列互补配对的mRNA。其调控的机制是通过互补配对而沉默相应靶位基因的表达,所以是一种典型的负调控机制。siRNA识别靶序列是有高度特异性的,因为降解首先在相对于siRNA来说的中央位置发生,所以这些中央的碱基位点就显得极为重要,一旦发生错配就会严重抑制RNAi的效应。
RNA干扰技术的前景
RNA干扰技术不仅是研究基因功能的一种强大工具,不久的未来,这种技术也许能用来直接从源头上让致病基因“沉默”,以治疗癌症甚至艾滋病,在农业上也将大有可为。从这个角度来说,“沉默”真的是金。美国哈佛医学院研究人员已用动物实验表明,利用RNA干扰技术可治愈实验鼠的肝炎。
尽管尚有一些难题阻碍着RNA干扰技术的发展,但科学界普遍对这一新兴的生物工程技术寄予厚望。这也是诺贝尔奖评审委员会为什么不坚持研究成果要经过数十年实践验证的“惯例”,而破格为法尔和梅洛颁奖的原因之一。
诺贝尔生理学或医学奖评审委员会主席戈兰・汉松说:“我们为一种基本机制的发现颁奖。这种机制已被全世界的科学家证明是正确的,是给它发个诺贝尔奖的时候了。”
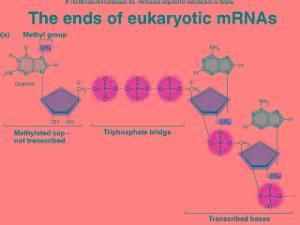
核糖核酸_核糖核酸 -转录
转录是指DNA的双链解开,使RNA聚合酶可依照DNA上的碱基序列合成相对应之信使RNA(mRNA)的过程. 在人体需要酵素或是蛋白质时,都会需要进行此过程,才能借由信使mRNA,将密码子带出核模外. 好让核糖体进一步的利用信使RNA(mRNA)来翻译,合成所需之蛋白质? DNA的碱基有A(腺嘌呤)、G(鸟嘌呤)、C(胞嘧啶)、T(胸腺嘧啶),而RNA之碱基无T(胸腺嘧啶), 取而代之的是U(尿嘧啶),也就是有A(腺嘌呤)、G(鸟嘌呤)、C(胞嘧啶)、U(尿嘧啶). 在DNA中,A与T以两条氢键连结,G与C以三条氢键连结,但RNA只有U而无T, 所以在转录时DNA上的若是A,mRNA就会是U,也就是取代原本T的位置? 如下图所示,右边DNA的一股碱基序列若为‘AAACCG’,而左方的DNA因配对而就会成‘TTTGGC’, 但因RNA无T这个碱基,只有U,因此合成出来的mRNA对应之序列就为‘UUUGGC’ 因为DNA太大,无法出入核膜(细胞核的膜),所以才需要有mRNA的出现,让mRNA可穿过核孔(核膜上的孔洞) 到达细胞质进行翻译(核糖体合成蛋白质的过程),因此,转录对不管是人类还是动物甚至是细菌 都是不可或缺的重要反应?
核糖核酸_核糖核酸 -翻译
游离在细胞质中的各种氨基酸,就以mRNA为模版合成具有一定氨基酸顺序的蛋白质,这一过程叫翻译
首先氨基酸与tRNA结合生成氨酰-tRNA
然后是多肽链的起始:mRNA从核到胞质,在起始因子和Mg 的作用下,小亚基与mRNA的起始部位结合,甲硫氨酰(蛋氨酸)―tRNA的反密码子,识别mRNA上的起始密码AuG(mRNA)互补结合,接着大亚基也结合上去,核糖体上一次可容纳二个密码子。(原核生物中为甲酰甲硫氨酰)
再是多肽链的延长:第二个密码对应的氨酰基―tRNA进入核糖体的A位,也称受位,密码与反密码的氢键,互补结合。在大亚基上的多肽链转移酶(转肽酶)作用下,供位(P位)的tRNA携带的氨基酸转移到A位的氨基酸后并与之形成肽键(―CO-NH―),tRNA脱离P位并离开P位,重新进入胞质,同时,核糖体沿mRNA往前移动,新的密码又处于核糖体的A位,与之对应的新氨基酰-tRNA又入A位,转肽键把二肽挂于此氨基酸后形成三肽,ribosome又往前移动,由此渐进渐进,如此反复循环,就使mRNA上的核苷酸顺序转变为氨基酸的排列顺序。
最后是多肽链的终止与释放:肽链的延长不是无限止的。当mRNA上出现终止密码时(UGA、U氨基酸和UGA),就无对应的氨基酸运入核糖体,肽链的合成停止,而被终止因子识别,进入A位,抑制转肽酶作用,使多肽链与tRNA之间水解脱下,顺着大亚基中央管全部释放出,离开核糖体。同时大小亚基与mRNA分离,可再与mRNA起始密码处结合,也可游离于胞质中或被降解,mRNA也可被降解。
核糖核酸_核糖核酸 -自身调控
2013年5月新一期《科学》杂志上报告说,英国约翰・英尼斯中心的研究人员发现的这种核糖核酸名为COOLAIR,是一种反义长链非编码核糖核酸。研究人员以模式植物拟南芥作为研究对象,通过遗传筛选和基因克隆等手段,发现COOLAIR受到一种叫做R环的特殊结构的影响。R环是由一条脱氧核糖核酸(DNA)与核糖核酸杂合链以及一条单链DNA所形成的特殊基因组结构,一般在基因表达转录核糖核酸时可以形成瞬时的R环,但很快会被去除。而他们观察发现,R环能够通过抑制COOLAIR发挥作用,从而让拟南芥提前开花。