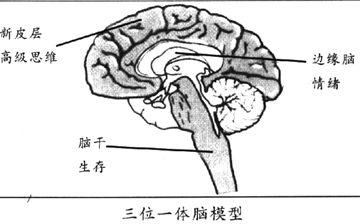
边缘系统是指高等脊椎动物中枢神经系统中由古皮层、旧皮层演化成的大脑组织以及和这些组织有密切联系的神经结构和核团的总称 。古皮层和旧皮层是被新皮层分隔开的基础结构。边缘系统的重要组成包括,海马结构、海马旁回及内嗅区、齿状回、扣带回、乳头体以及杏仁核。上述结构通过帕帕兹环[Papez 环路]相互联系,并与其他脑结构(新皮层、丘脑、脑干)有广泛联系,所以边缘系统的作用是使中脑、间脑和新皮层结构之间发生信息交换。 通过与下丘脑及植物神经系统的联系,边缘系统参与调解本能和情感行为,其作用是自身生存和物种延续。此外,海马结构还对学习过程和记忆发挥着突出的作用。因此如果海马结构或与之功能联系的结构受损,则导致遗忘综合征。其病变部位不同,产生的记忆障碍形式也不同。
边缘系统_边缘系统 -简介
边缘系统
边缘系统
limbic system
“边缘”一词源于拉丁语“limbus”。1878年法国解剖学家P.布罗卡提出“大边缘叶”的概念,用以指扣带回、海马回及其附近与嗅觉功能有关的大脑皮层。1937年J.W.帕佩茨提出,在组织学上从海马到乳头体,经丘脑前核、扣带回再返回海马构成了“边缘环路”,这个环路与协调情绪等高级功能有关。帕佩茨的理论引起了学者们的重视,也推动了以后的研究工作。由于环路内的联系复杂,密切,P.D.麦克莱恩于1952年进一步提出“边缘系统”这个概念。根据纤维之间的联系与功能特征, 在边缘叶基础上增添1. 额叶眶回、脑岛和颞极; 2. 皮质下核团: 杏仁核、隔核、上丘脑、下丘脑、丘脑前核及背内侧核一部分; 3. 边缘中脑, 指中脑被盖区的一些核团, 中央上核、脚间核等新结构, 称为边缘系统。对此已加以修正并扩充, 大致分为三个部分: 1.颞叶内侧边缘系统结构, 包括海马结构、杏仁体、扣带回和嗅周皮质( 而嗅脑则指大脑半球中接受与整合嗅觉冲动的皮质部分, 主要包括嗅球、嗅束、嗅三角、前穿质、杏仁体和海马旁回前部等) ;2. 丘脑内侧核团, 有内侧背核和前部核团; 3. 额叶的腹内侧部分, 包括眶额皮质、前额叶内侧.
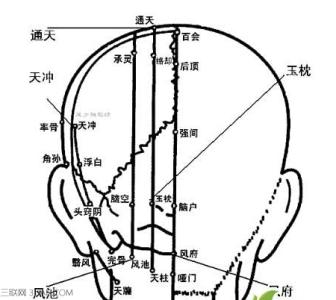
在大脑半球内侧面有一由扣带回、海马旁回及海马回钩等在大脑与间脑交接处的边缘连接成一体,故称边缘叶。边缘叶与邻近皮质(额叶眶部、岛叶、颞极、海马及齿状回等)以及与它联系密切的皮质下结构(包括与扣带回前端相连的隔区、杏仁复合体、下丘脑、上丘脑、丘脑前核、部分丘脑背侧核以及中脑内侧被盖区等)在结构与功能上相互间都有密切的联系,从而构成一个功能系统,称为边缘系统。
边缘系统_边缘系统 -功能
边缘系统的功能主要有以下几方面:
调节内脏活动
刺激边缘系统的后眶回、扣带回、岛叶、颞极、梨状皮层、旁杏仁皮层、后海马皮层等部位,可以引起人及动物的呼吸、血管以及其他内脏反应。刺激下丘脑不同部分所引起的内脏反应最为明显,在出现竖毛、瞳孔扩大的同时,血压急剧升高,心率加快以及出现饮水、摄食、排尿、排粪、流涎和呕吐等反应。刺激眶回皮层,可引起血压下降,心率变慢。一般认为,边缘系统对于心血管活动的影响是通过下丘脑和脑干低级心血管反射活动中枢而实现的。此外,边缘系统还可以通过下丘脑-垂体系统的所谓神经体液途径,影响下丘脑各种神经分泌,从而影响相应垂体激素的分泌,导致内脏功能活动的改变。实验证明,边缘系统中许多部位接受内脏传入神经发来的冲动,这种冲动对于边缘系统反馈性地调节内脏活动具有重要意义。已经发现,边缘系统中有一些神经元本身即是某种极为敏感的感受器,例如下丘脑部分有感受温度变化的神经元和感受血液内葡萄糖浓度变化的神经元。这些神经元的活动对于调节体温变化、消化液的分泌量以及进食活动都具有十分重要的生理意义。
调节中枢神经系统内的感觉信息
在低等脊椎动物,大脑的海马结构能够接受各种感觉刺激的影响。在高等哺乳动物,躯体、听觉以及视觉等感觉冲动能够传入海马;刺激边缘系统的下丘脑前区、扣带回等部位可以使痛阈升高;刺激杏仁核群能够使丘脑内膝状体的听觉信息受到阻抑。
影响或产生情绪
损伤猴、猫、狗等动物的杏仁前核、海马、视交叉前区、穹窿、嗅结节及隔区,可使动物出现“假怒”反应或“愤怒的行为”。也有研究证明,如果只将扣带回损坏而不伤及大脑新皮层,常使动物的情绪反应减弱或不易出现。发怒的阈值升高,出现一种“社会性的淡漠”或是“失却恐惧”的症状。这时,动物对于平常必须躲避的有害刺激,表现得无动于衷。
切除猫的杏仁核之后,出现与情绪反应有关的性功能亢进、性反应增强。切除猕猴的杏仁核,可使其行为在群居生活中由统治者的地位变为从属者的地位。临床研究表明,损伤边缘系统较为广泛的区域之后,病人极易发怒,在社交场合表现出强烈的情绪反应。这和利用动物所获得的实验结果也很相近。
曾有学者认为,调节“愤怒”情绪活动的主要神经结构位于下丘脑。切除动物丘脑而保存下丘脑,使动物出现“情绪呆板”,一触即怒,或是表现出挣扎、露爪、竖毛、瞳孔扩张、怒叫等明显的情绪反应。在脑内埋藏电极并刺激下丘脑的不同部位,可使动物出现攻击、发怒的行为和逃避或状如恐惧的行为。临床病例表明,双侧下丘脑腹内侧核受到肿瘤侵犯之后,病人经常出现攻击性行为。
研究证明,用电流刺激猫的边缘中脑区,会引起怒叫和攻击等情绪反应。位于滑车神经核平面的中脑外侧被盖区也被称之为怒叫中枢。神经解剖学的研究也证明,与情绪反应有关的下丘脑部分,有神经纤维发到边缘中脑区,因而刺激下丘脑所产生的种种反应,可能是通过边缘中脑区而起作用的。
研究者曾将电极埋在下丘脑以及边缘系统的其他部位,而将控制刺激开关的开关安装在实验箱内,使动物能够自己操纵,进行“自我刺激”。结果发现,动物的这种自我刺激有时每小时竟达5000次之多,如果不加干预,它可能持续到出现衰竭。引起自我刺激最有效的区域为下丘脑后部,即乳头体前区,其次为中脑被盖部分、隔区、内侧前脑束等边缘系统部位。
虽然边缘系统中许多部位的活动都能影响或产生情绪反应,但就整个系统而言,则难以定出某种情绪反应活动的中枢代表区的严格位置。在大多数情况下,各种情绪代表区在边缘系统内部有广泛的重叠范围。
引起睡眠活动
边缘系统中的后眶回、副嗅皮层、视前区以及下丘脑前部是与睡眠活动有关的部位,这些部位曾被一些学者统称为基底前脑区。用电流刺激这一脑区,动物出现睡眠反应。在这一脑区以下水平作切割手术,可以消除大鼠的失眠。临床观察证明,由于脑外科手术损伤基底前脑区的病人,也同样出现严重的失眠症状。
参与学习和记忆活动
很早以前,临床病例即证明,病员隔区损伤之后,便难以用概性的言语表达事物特征。损伤杏仁核之后,病人的应变能力减弱。海马与乳头体受到损伤,可以导致一种极为明显的记忆障碍,即过去经验保持的情况下,短期记忆丧失。自60年代关于边缘系统在学习和记忆中的作用有大量的动物实验研究的文献报道。损毁双侧海马之后,虽然能使动物建立操作式条件反射,和形成对不同图形的鉴别反射活动,但要求训练的次数大大增加。它们更难以建立以时间间隔作为条件刺激的反射活动,也无法培养条件性的延迟反射。已经具有延迟反射活动的动物,如果切除其海马,延迟反射也不易出现,但是仍旧保存其他条件反射。
海马受损之后,动物对周围环境中新异刺激的朝向反应增强。当新异刺激重复出现时,这种反应难以消退。这说明动物的“记忆”能力有损伤。但是对于已经建立的条件反射,在海马损毁之后并不消失,这说明海马不是保存过去经验痕迹的部位。很可能在学习过程中,各种刺激信息在海马留下暂时的痕迹,经过它的活动,刺激信息进入长时记忆。
从分子和细胞的水平,研究也证明海马结构参与记忆的形成。例如,实验证明,在学习过程中,动物的边缘系统中的一些部位核糖核酸的含量增高。利用放射自显影技术进一步证明,在鉴别亮度的条件反射活动过程中,海马的锥体细胞、扣带回神经元以及视区大脑皮层神经元内的尿嘧啶核苷酸明显增加,海马神经元的蛋白质合成率升高,细胞内核糖体数目也在训练过程中增加。神经组织学的研究证明,在学习初期海马结构中突触的数量增加,突触后膜的致密度增大,乙酰胆碱的含量增高。这些变化都说明海马结构神经元的活动增强,这对短时记忆和长时记忆的形成可能都十分重要。
边缘系统_边缘系统 -展望探索
由于边缘系统的所属部分的多种重要的生理功能,因而引起神经生理学、神经病理学、精神病学、心理学等学科工作者们的重视。重要的研究工作主要集中在以下几个方面:边缘系统中的神经递质以及多肽类化合物;边缘系统对于运动及行为的调节;边缘系统对于感觉功能的影响;边缘系统和学习及记忆功能。研究工作细至在分子水平上阐明机制,大则在动物行为方面进行整体或群体的考察。在研究方法上,充分利用新技术的同时,不拘一格地将多种研究方法综合使用,因而在许多研究工作中,出现将高分辨率的电子显微镜、微电泳、放射自显影、荧光组织化学、免疫组织化学、细胞内记录的电生理学方法以及条件反射或是操作性条件反射相互配合使用的情况。这种综合性的研究,更利于阐明这一系统的功能。当前一般认为边缘系统的主要功能,是对动物体的感觉、运动和内环境稳定等各种生理功能起调节作用。损伤边缘系统的任何一部,并不导致机体某种基本功能的丧失,而仅对某些刺激的反应中有些因素调节失灵。对于中枢神经系统,除了感觉、运动以及调节系统而外,增加边缘系统参与整合运动,可以使机体更易对复杂而多变的环境作出正确的反应。