缝隙连接是将相邻细胞的细胞质进行连接的细胞内通道。细胞间的通讯方式可分为间接与直接方式。以体循环远程分泌、旁分泌或自分泌方式经第二信使途径完成一系列生理、生化功能的调节方式称为间接通讯;而以细胞间的缝隙连接(gap junction)为途径进行的细胞间直接的信息交流,称为直接通讯,又称缝隙连接细胞间通讯(gap junction inter-cellular communication/GJIC)。
缝隙连接_缝隙连接 -概述
细胞间呈线状缝隙连接
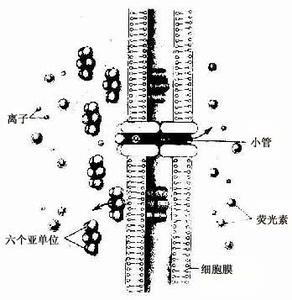
缝隙连接分布广泛,几乎存在于所有的动物细胞中。用超薄切片术可显示相邻两细胞的连接处的细胞质膜明暗相间七层结构,细胞间的缝隙约2纳米,其内有间隔的均匀排列的颗粒。用冰冻断裂电镜技术显示缝隙连接的颗粒区面积大小不等,且排列规则而密集。用X线衍射技术证明,每个颗粒由6个蛋白质亚单位构成,它们呈环行排列,中间有直径2纳米左右的小孔,被称为连接小体(con-nexon)。每两个连接小体相对合,并分别包埋在相邻细胞的质膜中,构成两个细胞之间的通道。连接小体成簇状排列,以增加通道的数量。通道只允许分子量小于1200的物质自由通过,如无机离子,氨基酸,葡萄糖等。缝隙连接是一种动态结构,有多种因素参与调节通道的开放和关闭,如细胞内pH、Ca2+浓度和细胞膜电位等。缝隙连接有多种功能,它与细胞的代谢和分化,物质的运输和电兴奋的传导等有密切关系。
缝隙连接_缝隙连接 -缝隙连接的组成与结构
组成
缝隙连接由相邻细胞膜上的两个连接子(connexon)相互锚定组成,而连接子是一个六聚体,由六个亚单位-连接蛋白(connexin/Cx)组成。连接蛋白是由十余个成员组成的一个保守大家族,分子量由26~56KD(≈2.6×104~5.6×104u)不等,可分为α-Cx和β-Cx.在啮齿类动物中,至少由13个基因编码,与其它种属有较高的同源性,各亚成员之间有50%~60%同源,主要的差别在于Cx分子的胞浆部分。
结构
连接蛋白的共同结构示意图
连接蛋白的共同结构如图所示,四个跨膜亲水片段称为M1-4,为α螺旋结构,两个胞外环(Extracellular loop)分别称为E1-2,和一个胞浆环(cytoplastic loop/CL)。连接蛋白的羧基末端与氨基末端位于胞浆内,氨基末端相对比较保守,而羧基末端则差别较大。羧基末端的丝/苏及酪氨酸残基的磷酸化/去磷酸化水平影响着缝隙连接的形成及功能状态,并能够感受胞内的信息而改变构象,从而调节缝隙连接的形成及传导性。胞外的E1与E2区各含三个半胱氨酸残基,E1区的三个半胱氨酸分别由1个和3个氨基酸残基分隔,E2区的半胱氨酸分别由4个与5个氨基酸残基分隔,但在Cx31中,后者分别由5个与5个氨基酸残基分隔,在E1环与E2环之间至少有一个二硫键连接。利用位点突变技术表明,在Xenopus卵母细胞中,Cx32的E1、E2区的任一半胱氨酸残基突变都导致缝隙连接的传导性的完全阻断,E1、E2区的半胱氨酸残基单个突变导致缝隙连接功能丧失,而成对的突变或第三位半胱氨酸残基的单个突变则不影响功能。而E1与E2的第一与第三位半胱氨酸互换,则传导性最高。E区形成堆栈的β-Sheet构象,E1、E2的反平行的β-Sheet结构与4个跨膜的α螺旋的顺序排列相关。
缝隙连接模型
连接蛋白可寡聚成六聚体即连接子或称半通道(hemichannel)。连接子可由单一连接蛋白组成称同聚体连接子(homometic connexon),也可有几种连接蛋白构成异聚体连接子(heterometic connexon),如在羊晶状体上皮间存在Cx46与Cx50的异聚体连接子。不同组合的异聚体连接子对物质的通透性具有选择性,而E2区域的相互作用与选择性通透有关。
由异聚体连接子参与组成的缝隙连接称作异型缝隙连接(heterotypic gap junction),反之称作同型缝隙连接(homotypic gap junction),在缝隙连接间的细胞间隙有35埃。
缝隙连接_缝隙连接 -缝隙连接的功能
缝隙连接的亲水孔道约1.5nm,允许分子量小于1.5 KD(≈0.15×104u)的离子、代谢物、及一些第二信使(cAMP、Ca2+、IP3)通过,通过物质交换构成GJIC。缝隙连接具有传导快、低阻抗、延搁时间短的特点,主要的生理功能有以下几个方面。
协调细胞间活动的一致性
在心肌细胞中,它的活动具有全或无现象,细胞间收缩功能的协调是通过缝隙连接来完成的。利用Cx43基因的定标突变技术,发现突变型杂合子小鼠(Cx43-/+)较野生型(Cx43+/+)心室外膜的电传导速度降低30%~40%,QRS波复极时间明显延长。在体外培养的大鼠心肌细胞中,随着细胞间的缝隙连接增多,细胞由不规则收缩趋向同步收缩。
参与信息的传递及神经冲动的传导
蓝色区域是缝隙连接
在一些细胞中,由于缝隙连接对细胞内第二信使如Ca2+的通透性,它能介导细胞间的信息传递。Toyofuku构建了一个模型,能同时表达Cx43与兰尼啶受体(一种胞内内质网上的Ca2+通道)的A细胞四周围以仅能表达Cx43的B细胞,从而形成缝隙连接,经机械刺激后,发现Ca2+经缝隙连接由A细胞向B细胞扩散。来自胚胎小鼠的星型细胞,富含由Cx43组成的缝隙连接,利用位点突变技术产生的纯合子(Cx43-/-)和杂合子(Cx43-/+)小鼠星型细胞,机械刺激汇合的星型细胞,发现由此触发的细胞内Ca2+上升的细胞数少于培养的野生型星型细胞。同样Ca2+的经缝隙连接传导在成骨细胞、软骨细胞、小鼠视网膜色素上皮细胞、胰岛素分泌细胞、羊的晶状体上皮细胞中存在。
参与细胞的分化生长与发育
同样利用Cx43缺陷的小鼠星型细胞,利用神经胶质细胞分化的标记物如神经胶质纤维原酸性蛋白和S100染色标记发现,在纯合子、杂合子及野生型星型细胞的分化都正常,但前两者的生长速度则减慢。Cx43-/-纯合子小鼠出生不久即死亡。在Clone9肝细胞的增殖中,由PKC导致磷酸化使缝隙连接失偶联是在G0/S期发生事件中必需的[9]。在肿瘤发生中,GJIC中断在肿瘤细胞的生长、转移中起重要作用,在肿瘤组织中,缝隙连接的数目明显减少,体外把肿瘤细胞与表达正常Cx43的正常组织细胞共同培养,则肿瘤细胞向正常分化。在生长发育过程中,一种Cx的表达可诱导或抑制其它Cx的表达,从而诱导细胞的正常分化生长。在胚胎期小鼠心脏发生中,准确的缝隙连接调节是保证心脏正常发生的关键,缺少Cx43基因的新生小鼠心脏发生异常。
缝隙连接_缝隙连接 -缝隙连接和心血管疾病
在心血管系统中,心肌细胞及平滑肌细胞间存在着丰富的缝隙连接,其主要成分以Cx43为主,因此缝隙连接的功能失常将引起各种心血管疾病。
心律失常
心脏电复极的不一致是导致心律失常的一个重要原因。心肌纤维化常伴有较高的室性心律失常,其最明显的病理结果是将心肌细胞隔离开来,从而失去缝隙连接的连接,失去电复极的一致性,为折返的形成创造了条件。在有突变型的杂合子(Cx43-/+)的小鼠,心肌外膜的电位记录显示心脏电传导降低30%~40%,而单个细胞的动作电位参数在突变型与野生型之间无区别。Shaw研究发现,当细胞间缝隙连接下降时,心肌单一传导阻滞的易损窗时间(vulnerable window time;VWt)及易损窗电位(VWpot)分别增加4.7倍及3.6倍,从而更易出现单向传导阻滞,引起折返。另外,心肌细胞的失偶联比心肌细胞的兴奋性降低更显著引起VWt及VWpot的延长。一种人工合成的抗心律失常肽AAP10(antiarrhymic peptide),是通过增加缝隙连接的传导性而起作用。在人类心脏组织的房室结、窦房结、及传导束中,含有丰富的Cx40,各传导纤维间的缝隙连接与激动的顺利传导密切相关。利用野生染色体连锁分析,位于1号染色体1p1-1q1区间的Cx40的突变,是造成一种常染色体显性遗传的传导阻滞的候选基因。
先天性心脏病
某些先天性心脏病的发生于缝隙连接的突变或功能丧失密切相关,这种突变可以是羧基末端的缺失,或磷酸化位点的点突变为主,从而影响Cx的功能。如上所述,一种以传导阻滞及扩张性心肌病(DCM)为临床表现的罕见先天性常染色体显性遗传性心脏病可能系Cx40的突变造成的。而当Cx43过度表达时,则形成小鼠的心脏及神经管的缺损。在另一种罕见的心脏异位伴侧壁缺损的先天性心脏病中,发现存在S364位突变成脯氨酸,从而干扰了羧基末端的正常磷酸化调节及其受PH的调节。
缺血性心脏病
心肌缺血及心肌梗死后,心律失常是最常见并且是潜在的具有生命危险的并发症。除了心肌纤维化以及低氧可以引起缝隙连接的功能障碍外,其它的具体机制尚未明了。正常的缝隙连接被限制于闰盘周围,犬心肌梗死模型的心肌免疫组化研究发现,心肌梗死区的尚存活但有变性的心肌细胞间,原先的缝隙连接发生重构,缝隙连接沿侧壁垂直族集排列,为室性心律失常形成折返环路。而远离心肌梗死部位的心肌组织的缝隙连接分布正常,但缝隙连接表面积/细胞体积比下降了47%,缝隙连接数目/细胞比下降了30%。在心肌梗死愈合区的边界的心肌细胞的电传导降低,并且缝隙连接的组织方式明显不规则,缝隙连接已不限于闰盘,而广泛地分布于细胞表面。
心肌病
Cx40的突变被认为是形成一种遗传性扩张型心肌病(DCM)的一个候选基因。而Cx40主要位于传导系统中,故其突变如何引起DCM的机制尚待进一步研究。在肥厚型心肌病(HCM)中,在心脏细胞排列紊乱区,闰盘排列也紊乱失去规则,缝隙连接在心肌表面随机分布,而不局限于闰盘。细胞间缝隙连接数目增多,缝隙连接形状发生异常,以及分布重构是形成心律失常的基础,也就解释了伴随HCM的心律失常产生与维持的原因。
感染性心脏病
Chagas病,一种南美洲锥虫病,在拉丁美洲是引起心功能不全及心律失常的一个主要病因,其病程可以表现为急性及慢性过程。在啮齿类动物的心肌细胞培养时,给予急性感染,发现心肌细胞的同步收缩变得不规则,细胞间的缝隙连接减少,心肌细胞的电生理特性改变,动作电位时程变短,胞内Ca2+增高,α肾上腺素敏感性降低。来源于慢性感染兔的血浆经Langendorff兔心脏灌流时发现有心电图异常,细胞间Lucifer Yellow染料的交换速度与程度明显减少。因此,可能激素及体液免疫也影响着缝隙连接的功能。
动脉粥样硬化
动脉粥样硬化的病理生理本质是平滑肌细胞(DMC)的增生,及细胞外基质的过度合成。过去所重视的多是各种细胞,经细胞因子作用,经第二信使介导的间接细胞间通讯对SMC的作用,而对于SMC间的缝隙连接对动脉粥样硬化发展的作用则研究较少。
目前认为,动脉粥样硬化的发生被认为是一个类似肿瘤发生的一种良性过程。既然在肿瘤发生中,GJIC的中断是必需的,那么,缝隙连接的功能及数目的改变必然与动脉粥样硬化的发生密切相关。
正常的血管组织中,完整的内皮细胞是维持血管正常生理功能是必需的,而Cx43是维持内皮连续性及完整性所必需的。在动脉粥样硬化发生中,存在着内皮细胞的障碍,如内皮通透性增高,内皮的不连续;另外,内皮能与循环中的白细胞经缝隙连接而诱导白细胞的趋化,促其分泌细胞因子及化学物质,从而介导动脉粥样硬化的形成。在脂多糖处理的兔缺血再灌注模型中,淋巴细胞表达Cx43,并与内皮形成缝隙连接。血液中的各种内皮毒性物质(如LDL、胰岛素、高葡萄糖)也可破坏内皮间的缝隙连接形成。而同样失去完整的缝隙连接,NO诱导的血管舒张作用便失去了作用。
SMC的转型(由收缩型向分泌型转变)及增生,向内膜移行及吞噬脂质是动脉粥样硬化发生的一个关键。在此过程中,正常的位于中膜的SMC,必定要摆脱缝隙连接对细胞的固定作用。利用免疫组化的研究表明,在动脉粥样硬化发生的早期,及内膜增厚期及早期斑块中,荧光斑点数目增多,但直径变小,随着病变进展,荧光斑点直径变大,但数目变少。但在一些丝裂原物质的刺激下,通过Cx43的磷酸化,缝隙连接的功能是抑制的,同样SMC的体外培养表明,细胞间缝隙连接是抑制SMC增殖的。在由收缩型向分泌型转变后,SMC的Cx43表达增多达6倍,缝隙连接的直径也变大了。在大鼠肝上皮细胞间,经血小板源性生长因子(PDGF)刺激后,缝隙连接解体,Cx族集成块状,体积变大,最终由溶酶体吞噬。既然在丝裂原刺激下,缝隙连接是受抑制的,那与动脉粥样硬化早期病变中,荧光标记的增多是相互矛盾的。因此推测,在丝裂原物质的刺激下,Cx43被磷酸化,使原先的缝隙连接解体,从而形成两个半通道,抑制或干扰Cx43的正常转运及装配,使Cx滞留于胞浆内,因而免疫组化时荧光染色数目增多,但颗粒直径变小;到了晚期,Cx43逐渐族集成斑块状,经类似于受体介导的吞噬作用,被溶酶体吞噬而降解,故荧光染色数目减少,颗粒直径变大。陈构建了双细胞的立体网状模型来研究缝隙连接,发现族集成斑块状的缝隙连接是很少具有通讯功能的。
循环中的单核/巨噬细胞不表达Cx43mRNA。然而原位杂交表明,来源于动脉粥样硬化病变区的单核/巨噬细胞来源的泡沫细胞则表达丰富的Cx43mRNA。在动脉粥样硬化发生过程中,单核/巨噬细胞经VLA-4与内皮细胞的VCAM-1结合而粘附,从而诱导缝隙连接Cx43的表达。由此推测,缝隙连接的形成,可能与单核/巨噬细胞的锚定、趋化渗透及吞噬脂质有关。
缝隙连接_缝隙连接 -缝隙连接的形成机理的新发现
最近,Shaw等在Cell上发表文章表明,连接蛋白快速、直接靶定在细胞粘附接触面的机制。这个过程涉及到微管,微管正极示踪蛋白(plus-end tracking protein, +TIP) EB1和与EB1相互作用的蛋白p150(Glued)将微管限制在粘附面,如同细胞-细胞交界面的皮层锚。封面描述的是两个邻近的心肌细胞,在细胞-细胞接触点上发展管缝连接片层。被限定的微管形成的管缝连接hemichannels导致形成片基和细胞质交流。
细胞连接
长期以来,人们一直认为gap junction的形成过程,第一步是hemichannels径向扩散(lateral diffusion),随后hemichannels由微管介导传递系统到达浆膜后形成gap junction。
加州大学医学院心血管研究实验室等部门的研究人员发现细胞以一种依赖微管的途径,通过粘附连接蛋白N-cadherin和β-catenin、微管正极示踪蛋白(plus-end tracking protein, +TIP) EB1以及与EB1相互作用的蛋白p150(Glued),将hemichannels直接靶定到细胞-细胞连接处。
在活细胞显微方法,如光褪色后的荧光复活(fluorescence recovery after photobleaching ,FRAP)、全内反射(total internal reflection fluorescence ,TIRF)、去卷积(deconvolution)和siRNA敲除等方法的基础上,研究人员推测微管正极优先拴缚(preferential tethering)在粘合连接(Adherens junction),会促进将connexin hemichannels直接递送到细胞-细胞交接面。这些发现提供了一个蛋白靶向细胞-细胞接触点传递的新机制。