Ti是在根瘤土壤杆菌细胞中存在的一种染色体外自主复制的环形双链DNA分子。是英文肿瘤诱发(tumor-inducing)的缩略式。Ti质粒大约在160~240kB之间。农杆根瘤菌之所以会感染植物根部是因为植物根部损伤部位分泌出酚类物质乙酰丁香酮和羟基乙酰丁香酮,这些酚类物质可以诱导Vir(Virulenceregion)基因的启动表达,Vir基因的产物将Ti质粒上的一段T-DNA单链切下,而位于根瘤染色体上的操纵子基因产物则与单链T-DNA结合,形成复合物,转化植物根部细胞。首先在E.coli中筛选重组分子,然后将重组质粒转化到农杆菌中,质粒与Ti质粒上的同源序列发生同源重组,将外源基因整合到Ti质粒上,用于侵染植物细胞。
ti质粒_Ti质粒 -Ti质粒与TDNA的整合机制
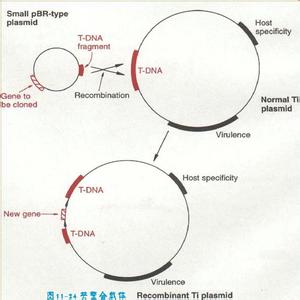
外源基因整合到T-DNA
几乎所有双子叶植物都容易受到土壤农杆菌感染,而产生根瘤。它是一种革兰氏阴性土壤杆菌(A.tumefaciens)。其致瘤特性是由Ti(tumor-inducing)质粒介导的。农杆根瘤菌之所以会感染植物根部是因为植物根部损伤部位分泌出酚类物质乙酰丁香酮和羟基乙酰丁香酮,这些酚类物质可以诱导Vir(Virulenceregion)基因的启动表达,Vir基因的产物将Ti质粒上的一段T-DNA单链切下,而位于根瘤染色体上的操纵子基因产物则与单链T-DNA结合,形成复合物,转化植物根部细胞。T-DNA上有三套基因,其中两套基因分别控制合成植物生长素与分裂素,促使植物创伤组织无限制地生长与分裂,形成冠瘿瘤。第三套基因合成冠瘿碱,冠瘿碱有四种类型:章鱼碱(octopine)、胭脂碱(nopaline)、农杆碱(agropine)、琥珀碱(succinamopine),使农杆菌生长必需的物质。
1.Ti质粒的结构
在发现根瘤农杆菌诱发冠瘿瘤的本质是Ti质粒后,Ti质粒便成为冠瘿瘤形成基因鉴定与分析的主要研究对象。
Ti质粒大约在160~240kB之间。其中T-DNA大约在15kb-30kb。Vir基因区在36kb左右。除此之外,Ti质粒上还存在Con区(regionencodingconjugation)和Ori区(originofreplication)。
c.Ti质粒约为200kb,重组操作非常苦难,也很难找到单一的酶切位点。
d.Ti质粒不能在大肠杆菌中复制,为了使重组质粒DNA的大量扩增,须添加入大肠杆菌复制子。加入植物细胞的筛选标记,如neor基因,使用植物细胞启动子及末端polyA化信号,加入多聚人工接头以利于外源基因的。
植物中一般不存在质粒,为利用农杆菌的Ti质粒,发展了共整合系统和双元载体系统,避免了在大的Ti质粒上进行分子重组操作的困难。
2.植物细胞转化的共整合系统
T-DNA在大肠杆菌质粒上,含有E.coli的选择标记和植物选择标记Kmr。首先在E.coli中筛选重组分子,然后将重组质粒转化到农杆菌中,质粒与Ti质粒上的同源序列发生同源重组,将外源基因整合到Ti质粒上,用于侵染植物细胞。T-DNA重组分子整合到植物细胞染色体DNA上,Kmr筛选转化细胞。
3.植物细胞转化的双元系统
目前T-DNA转化植物细胞的标准方法是双元系统,即穿梭质粒。插入外源基因的重组穿梭质粒直接转化含有Ti质粒的根瘤农杆菌,经筛选后直接感染植物细胞。与共整合系统所不同的是,含外源基因的质粒可在农杆菌内自主复制并保留下来。农杆菌侵染植物细胞后,植物的创伤信号启动Ti质粒上的Vir基因,随后将穿梭质粒的T-DNA切割下来,转移到植物细胞中。
T-DNA上共有三套基因和左右两个边界,LB和RB是长为25bp的末端反复重复顺序,在切除及整合过程具有重要意义。
tms由两个成:tms1(iaaM)和tms2(iaaH)
tmr由一个成iptz:
tmt由若干基因构成,合成稀有氨基酸衍生物,称为opines。它有三个成员:octopine=精氨酸与丙酮酸的缩合物,Napaline=精氨酸与-酮戊二酸的缩合物,Agropine=谷氨酸与二环糖的缩合物。
据此可将Ti质粒分为三大类,感染的植物诱导合成这些有机碱,但不能利用它们,其分解酶基因在Ti质粒上,分解产物为氨基酸和糖类,供根癌农杆菌使用作为氮源及碳源。
2.T-DNA的整合机制:T-DNA的详细整合机制尚不清楚,但有几个环节是明确的:
T-DNA切除由Vir区编码的特异性内切酶完成,分别在LB和RB的第三个碱基和第四个碱基之间产生缺口,并形成单链T-DNA。
T-DNA的LB和RB在整合中的作用是不对称的,RB顺序与整合有关,而LB无关。
T-DNA的整合可以是单拷贝的,也可以是多拷贝的,成串联形式排列,但整合位点的特异性尚未确定。