人禽流感,即人禽流行性感冒,是由禽甲型流感病毒某些亚型的毒株引起的急性呼吸道传染病。1997年5月,中国香港特别行政区1例3岁儿童死于不明原因的多脏器功能衰竭,同年8月经美国疾病预防和控制中心以及世界卫生组织(WHO)荷兰鹿特丹国家流感中心鉴定为禽甲型流感病毒H5N1引起的人类流感,这是世界上首次证实禽甲型流感病毒H5N1感染人类。之后相继有H9N2、H7N7亚型感染人类和H5N1再次感染人类的报道。
人禽流感_人禽流感 -病原学
人禽流感禽流感病毒属甲型流感病毒。甲型流感病毒呈多形性,其中球形直径80~120nm,有囊膜。基因组为分节段单股负链RNA。依据其外膜血凝素(H)和神经氨酸酶(N)蛋白抗原性的不同,目前可分为15个H亚型(H1~H15)和9个N亚型(N1~N9)。甲型流感病毒除感染人外,还可感染猪、马、海洋哺乳动物和禽类。感染人的禽流感病毒亚型主要为H5N1、H9N2、H7N7,其中感染H5N1的患者病情重,病死率高。
禽流感病毒对乙醚、氯仿、丙酮等有机溶剂均敏感。常用消毒剂容易将其灭活,如氧化剂、稀酸、十二烷基硫酸钠、卤素化合物(如漂白粉和碘剂)等都能迅速破坏其传染性。

禽流感病毒对热比较敏感,65℃加热30分钟或煮沸(100℃)2分钟以上可灭活。病毒在粪便中可存活1周,在水中可存活1个月,在PH<4.1的条件下也具有存活能力。病毒对低温抵抗力较强,在有甘油保护的情况下可保持活力1年以上。
病毒在直射阳光下40~48小时即可灭活,如果用紫外线直接照射,可迅速破坏其传染性。
人禽流感_人禽流感 -高致病性禽流感
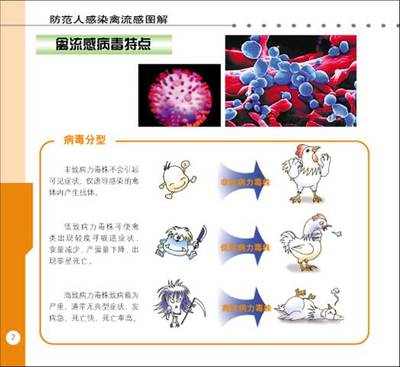
根据禽流感致病性的不同,可以将禽流感分为高致病性禽流感、低致病性禽流感和无致病性禽流感。由H5N1血清型引起的禽流感称高致病性禽流感,发病率和病死率都很高,危害巨大。
人禽流感_人禽流感 -传染源
传染源是指体内有病原体生存、繁殖并能排出病原体的动物和人。禽流感的传染源主要是病禽和带毒禽,包括水禽和飞禽,主要是鸡、鸭、鹅等家禽,特别是鸡。研究显示,对于高致病性禽流感,1克污染的粪便中病毒的含量就可造成100万只禽感染。目前还不能认为人类是禽流感的传染源。当然不排除其它禽类或猪成为传染源的可能。
人禽流感_人禽流感 -传播历史
1878年,Perroncito首次报道了在意大利鸡群中发生的家禽疫(fowl plague,禽瘟),直到1902年其病原才被分离出,这也是第一株被证实的流感病毒。而第一株人流感病毒直至1933年才被分离出。1941年,Hirst发现了流感病毒的血凝素活性。1955年,Schafer证实,家禽疫病毒为一种甲型流感病毒。对1957年和1968年人流感大流行病毒株起源的找寻促进了对动物中流感病毒生态学的广泛研究,在此期间从禽类中分离出了许多非致病性禽流感病毒,这才使人们意识到,禽流感病毒在禽类中分布是如此之广,在野生水鸟中更是普遍存在。此后,又在鸡和燕鸥中分离出了H5高致病株。
人们根据其毒力将禽流感病毒分为2型,高毒力性或称高致病性(可引起家禽疫)和无毒力性(仅引起轻症疾病或无症状感染)。少数情况下,实验室中的低致病性病毒也可在现实生活中导致禽流感暴发(如1978年美国明尼苏达州140个火鸡养殖场的禽流感暴发),但总的来说,其发病率和病死率比高致病性病毒低得多。
近30年来,高致病性禽流感病毒在下列地区的家禽中发生过暴发:澳大利亚[1976(H7)、1985(H7)、1992(H7)、1995(H7)和1997(H7)],英格兰[1979(H7)和1991(H5)],美国[1983-1984(H5)],爱尔兰[1983-1984(H5)],德国[1979(H7)],墨西哥[1994-1995(H5)],巴基斯坦[1995(H7)],意大利[1997(H5)],香港[1997(H5)],2003年10月以来亚洲多国的H5N1型禽流感和今年2月初美国的H7型禽流感。显而易见,所有的高致病性禽流感皆为H5或H7亚型,而N亚型似乎与毒力无关。此外,在严重的家禽疫样暴发中还曾分离出H4和H10亚型。
野禽――家禽
野禽――甲型流感病毒的天然宿主
已经在人、猪、马、海洋哺乳动物(如海豹和鲸)和禽类等多种动物体内分离出甲型流感病毒。种系发生研究不但揭示了甲型流感病毒的种特异性基因和跨种基因,而且证实感染其他物种的所有甲型流感病毒都来源于野生水禽。但流感病毒并不在野生水禽中引起致死性疾病,表明这种天然宿主已经在最大程度上适应了流感病毒。
大多数从野禽中分离出的禽流感病毒都是没有毒力的,即使某种禽流感病毒对一种禽类有高度致病性,对其他种类的禽类也可能不具有致病性。
人禽流感_人禽流感 -传播途径
迁徙的野禽可以长距离携带病毒(病毒在其肠道内进行复制),从粪便和其他分泌物中排出大量病毒,沿途感染家禽。被野禽污染同时又有家禽分布的水源是重要的传播链之一。水源性传播也为禽流感病毒年复一年的存在提供了可能。Ito等从阿拉斯加的湖泊中分离出了大量禽流感病毒,而这些湖泊正是许多候鸟的繁殖场所。
私藏或走私珍稀禽类(如斗鸡等)等人类行为也可导致禽流感病毒的地理性播散。
家禽之间的传播
禽流感很容易在同一个养殖场的家禽之间和不同的养殖场之间传播。由于病禽的呼吸道分泌物和粪便中含有大量病毒,饲料、饮水、设备和笼子等都可能受到污染,这就促进了病毒的扩散和在家禽之间的传播。污染的设备、运输工具、饲料、笼子或衣物(尤其是鞋子)可造成病毒在养殖场之间的传播。动物(如啮齿类)也可携带病毒,担当传播疾病的“机械性媒介”,目前还没有证据显示苍蝇也是机械性媒介。
禽类――哺乳类
传染到猪
迄今为止,已经从猪中分离出H1、H3、N1、N2和N7等各种亚型的流感病毒。
流感病毒从人传给猪也已有多次报告,且皆为H3亚型。在欧洲,禽H1N1病毒和人H3N2病毒自1979年起就共存于猪中,最终在猪中产生了一种重组病毒,其HA和NA基因为人H3N2病毒来源,其余部分为禽H1N1病毒来源。该病毒后来还感染了荷兰儿童,表明在猪中进行的禽与人流感病毒之间的遗传学重组,可导致有人类致病能力的杂交株的产生,目前循环于欧洲的猪H3N2病毒就是该重组病毒的后代。对于具有导致人类流感大流行可能的人-禽重组株的产生,猪可充当“混合器”的作用。1992年,Brown等从英格兰的猪中分离出了一种罕见亚型h1n7,其NA和M基因来源于马,其他来源于人。猪对禽类和哺乳类动物的许多流感病毒都易感,应将其作为检出具有大流行特性病毒的早期警报系统的一部分,进行定期监测。
Ito等发现了另外一个值得注意的现象:当禽H1N1病毒从禽传给猪后,病毒在猪体内复制时其受体特异性发生了改变。禽H1N1病毒识别NeuAcα2